
+91 6002993949
submission@iarconsortium.org
Open Access
ISSN (Print) : 2709-3301
ISSN (Online) : 2709-331X
The molecular mechanism of antibody-dependent enhancement in dengue virus infection is extremely complex and still, it is not clearly understood. In the past few decades, there were many cases of DHF/DSS with very high morbidity and mortality and a steady rise of new cases. Thus a clear understanding of the molecular pathogenesis of ADE is essential. In Dengue fever, DHF/DSS usually occurs by the subneutralizing or cross-reacting hetero-serotype antibodies acting on the Dengue virus. These antibodies enhance the entry of the virus via activating Fc gamma receptors FcγR. In absence of antibody, cell binding, cell entry, replication, assembly, secretion, antiviral responses are all quite different than the infection which occurs under influence of antibodies of a different serotype. In the molecular mechanism of ADE, it is important to know how interferons are suppressed and how the dual outcome of antigen-antibody reactions is important as it determines the fate of the infected person. A detailed description of all these issues is given in the manuscript.
More than 400 different viruses can infect humans. Human diseases caused by viruses are particularly interesting considering the small amount of genetic information introduced into a host cell. This apparent simplicity belies the severe pathological features and clinical consequences that result from many viral diseases. In general, only preventive or supportive treatment is available in viral diseases. Altogether, these diseases are one of the stereotypical and yet most perplexing of all infectious diseases. Approximately 150 of the recognized arboviruses caused illness in human beings. Diseases produced by the arboviruses can be divided into three clinical syndromes:
Fever of an undifferentiated type with or without rashes
Encephalitis often with a high fatality rate
Hemorrhagic fever, severe and fatal
No vaccines are available for most human arthropod-borne viral diseases, only supportive treatment is beneficial. An example of one of these types of viruses is the dengue virus. Dengue virus is an arthropod-borne virus or arbovirus, which is transmitted by blood-sucking arthropods, Aedes mosquito from one vertebrate to another. The virus proliferates in the arthropods’ tissue and stays lifelong without producing any disease.
Dengue fever often leads to hemorrhagic fever. Patients suffer from headaches, muscle pain, flushing of the skin, massive hemorrhages either locally or throughout the body, shock and death. Every year around 50-100 million people are infected with dengue fever in most tropical and subtropical regions and about 100 countries are at risk for dengue transmission [1,2]. Usually, different strains of the virus interacting with people with different immune backgrounds lead to complex outcomes. One of the reasons is cross serotype immune response, where a previously dengue-infected individual again gets infected for a second, third, or fourth time with other serotypes of the virus. This mechanism is known as Antibody-Dependent Enhancement (ADE). The previous antibody of the initial strain dengue virus now interferes with the immune response to the current strain, leading paradoxically to more virus entry and uptake [3].
In this article, we have discussed how this ADE complicates the dengue disease and what are the mechanisms behind it. Besides, the structure of the dengue virus and life cycle are also discussed herewith.
Structure of Dengue Virus
Dengue virus or DENV, a mosquito-borne virus has a roughly spherical structure. It is a single positive-strand RNA virus [+ssRNA] that belongs to the Flaviviridae family and genus Flavivirus [4].
The 11Kb ssRNA encodes for three structural proteins i.e capsid protein C, membrane protein M, envelope protein E and seven nonstructural proteins i.e NS1, NS2a, NS2b, NS3, NS4a, NS4b and NS5. Short non-coding regions are also included on both the 5’ and 3’ends. In the core of the virus, there is a nucleocapsid that is composed of the viral genome and capsid protein and outside a lipid bilayer membrane, called the viral envelope, surrounds the nucleocapsid [5].
The E protein or envelope is a dimeric protein that helps to attach the virus initially to the host cell. Three ectodomains, ED1 to ED3 and a transmembrane part complete each E protein monomer. ED2 includes the dimerization interface, two glycosylation sites and the peptide of fusion with the cellular membrane. ED3 is a compact and immunoglobulin-like continuous polypeptide segment. ICAM3-grabbing nonintegrin, CD209, Rab5, GRP78 and several other proteins interact with the viral E protein [6]. For serological detection and vaccine production, recombinant viral E protein is used [7].
The M protein or membrane protein are composed of seven anti-parallel β strands stabilized by the three-disulfide bond. It has a crucial function in the formation of the viral particle [6]. 180 copies of icosahedral structured E glycoprotein is present on the surface of immature and mature DENV particles. These E-proteins form a complex with M protein in the mature virus and turn to form dimers with other E-M complexes. With three of these dimers, it form a raft and 30 such rafts arrange in a herringbone pattern on the surface of the virus [8]. The transmembrane M protein possesses an extension at its N-terminus forming a globular β-barrel domain or pr domain, so it is called prM. The immature particle has a completely different structure from a mature virus at neutral pH. In the case of maturation, this pr domain caps the fusion loop of the E protein, preventing the newly synthesized immature virus from fusing back into the cell when moving through the acidic compartments of the trans-Golgi network [9].
The NS1, DENV nonstructural protein 1 is a 46 KD glycoprotein. It has several structural formations depending upon the place of secretion. It is expressed as a monomer; on the cell surface, it forms a membrane-bound dimer and into the blood circulation of patients, it is secreted as a hexamer consisting of three dimers of ~70 lipid molecules which forms an open barrel structure with a hydrophobic central cavity. This dimeric NS1 protein helps in viral genome replication by interacting with NS4A and NS4B transmembrane proteins in the early stages of infection [10]. In the case of a dengue fever diagnostic kit, NS1 possesses a pivotal role in the detection of the virus in patient sera [11]. Although NS1antigen may present in the blood up to 9 days’ post-infection; but test results up to 5 days post-infection is highly significant.
The DENV NS3 protein are a trypsin-like serine protease. In the N-terminal one-third of the protein, a serine protease catalytic triad i.e. histidine 51 (His51), aspartic acid 75 (Asp75) and serine 135 (Ser135) are accommodated within 180 amino acids. The catalytic site of the protease enzyme requires the central ~40 residue hydrophilic domain from NS2B to perform effectively (NS2BH-NS3pro) [12]. The protease then targets to cleave between the following sites i.e NS2A-NS2B, NS2B-NS3, NS3-NS4A and NS4B-NS5. It also targets the signal sequences present at the upstream of C-prM and NS4A-NS4B junctions, as well as NS2A and NS3 itself [13].
This NS3 protein has been found to exert other enzymatic activity during viral replication. The C-terminal rest of the two-third of the protein, contains a nucleoside triphosphatase (NTPase), a 5’ RNA-triphosphatase (RTP) and helicase. Due to its role in the survival of the virus, NS3 protein makes itself a good choice to be studied as a drug target [14].
The role of NS4 protein is yet to be understood. These transmembrane proteins NS4A and NS4B harbor an enzyme activity by acting as a scaffold for the replication complex formation. 2K, a transmembrane protein connects the two transmembrane proteins and then cleaves during the polypeptide maturation. The NS4A induces the localization of the endoplasmic reticulum and the replication complex for the perinuclear area stabilization. It is important due to the need for RNA viruses to use nuclear components for replication [15]. On the other hand, NS4B modulates viral replication by interacting with the helicase domain of NS3 to assist its dissociation from the viral RNA [16].
The largest and most conserved flaviviral protein, NS5 protein, consists of about 900 amino acids and has a molecular mass of about 100 kDa. It harbors an RNA-dependent RNA polymerase (RdRp) domain at its C-terminal end which is vital for viral replication. This protein also contains a capping site, made up of a methyl transferase (MTase) and guanyltransferasse (GTase) domain at its N terminus [17]. The protein is predominantly located in the nucleus of infected cells, to suppress the host antiviral response. The replicative role of the protein helps to decide the virus's longevity and survival, so it is a promising antiviral target [18].
Dengue Serotype and structural heterogeneity
The dengue virus has four related but antigenically distinct serotypes: DENV-1, DENV-2, DENV-3 and DENV-4. Each of these viruses generates a unique host immune response to the infection. They are genetically similar and share approximately 65% of their genomes [19]. Though it was believed that dengue infections are caused by four serotypes, but recently in 2013, another serotype, DENV -5 has been reported [20].
DENV shows structural heterogeneity also. Some DENV strains have been shown to undergo dramatic structural rearrangements from the compact smooth to bumpy surface particles after changing the incubation temperature from 28˚C to 37˚C. Cryo-EM reconstruction of the 37˚C of the DENV sample showed four structural classes with different radii representing the various stage of expansion [20]. The class I virus possesses the same compact smooth structure as a mature particle. In the case of class II and III particles, the envelope or E proteins remains intact except at the icosahedral twofold vertices. In class IV particles, it is smaller than other classes and the E protein layer is absent [21]. Due to the more solvent accessible of their E proteins, these expanded particles express different antigenic characteristics compared to the previously compact virus structure. It has been shown in in vitro studies that anti-prM antibodies and anti-fusion loop have a crucial role in ADE by binding to partially or fully immature viral particles [22]. As the E proteins have now more solvent-accessible, it will allow fusion loop antibodies to bind [21].
Dengue Virus Life Cycle
The dengue viral replication process begins when the virus attaches to a human skin cell via receptor-mediated endocytosis. After entry inside the cell, the viral particle first uncoated the nucleocapsid due to high exposure to the low pH of the endosome by inducing a large conformational change at the surface. After releasing the RNA molecule, it is translated as a single polyprotein [23]. While in this process, the signal- and stop-transfer sequences of the polyprotein direct it’s to and fro translocation across the Endoplasmic Reticulum (ER) membrane. Using the cellular and virus-derived protease; the polyprotein is processed into three structural proteins (C, prM and E) and seven Non-Structural (NS) proteins [NS1, NS2A, NS2B, NS3, NS4A, NS4B and NS5] via co- and post-translational modification. The glycosylation of amino acid residue Asn67 and Asn153 assure proper folding of E-protein [24]. The NS proteins begin to initiate viral genome replication after protein translation and folding. The C protein helps to form nucleocapsid by packaging newly synthesized RNA. Then the prM and E proteins are oriented into the lumen of the ER as heterodimers. The prM-E complexes on the newly synthesized particles are arranged as 60 trimeric spikes, which guides virion budding by inducing a curved surface lattice [25]. The maturation of immature particles formed in the ER by traveling through the secretory pathway. The slightly acidic pH [~5.8-6.0] of the Trans-Golgi Network (TGN) triggers dissociation of trimeric spikes into E protein dimers as it has been observed in a mature virus. This exposed the furin cleavage site on the prM to cleave pr from M and both of which attached to the viral surface due to the low pH environments. In the extracellular environment, the neutral pH causes the pr protein to dissociate from the virus particle upon secretion. Then a fully mature virus is formed by uncapping the E protein fusion loop [26].
Immune Response and Antibody-Dependent Enhancement
After the introduction of the DENV virus into the skin via female mosquito, DENV is challenged by the host's innate immune system by producing interferons (IFNs). The production of type 1 IFNs starts after the virus infects interstitial dendritic cells within an hour. In in vivo and in vitro studies, it has been found that both type I (α,β) and type II (γ) interferons are crucial against DENV infection [27]. Interaction of virus with Pathogen Recognition Receptors (PRRs) initiates IFN production. C type lectins (DC-SIGN, MR, CLEC5) and toll-like receptor (TLR3, TLR7) are these types of PRR which convey their signals through several transcription factors which ultimately induce the expression of IFN. Further, it activates the JAK/STAT pathway to express more than 100 effector proteins [28]. Interestingly, it has been found that DENV also suppresses type I IFN response. The viral NS proteins (NS2A, NS4A, NS4B and NS5) block the IFN signaling by reducing STAT activation. This is mediated by both non suppressive and suppressive strains that block STAT1/STAT2 pathways [29].
Not only innate immunity, but the humoral immune response also develops approximately 6 days after a bite from DENV infected mosquito against the E and prM glycoproteins present on the surface of the virus (Figure 1). Antibodies are also generated against NS1 antigens [30]. Antibodies play a dual role in controlling DENV infection as it has been found to both neutralize and enhance DENV infectivity in vivo and in vitro [22]. The neutralization of DENV occurs when several antibodies docked on the virion exceeds a certain threshold [31]. These neutralizing antibodies are strain-specific and created against domain III of the E protein but in the case of weaker and cross-reactive neutralizing antibodies, it is usually located to domain II or within the fusion peptide [32].
But sometimes, some cells of the immune system lack the usual receptor on their surface that the virus uses to gain entry, but they have Fc receptors that bind to one end of the antibodies. This results in higher virus burden and eventually enhancement of disease. This is observed usually in case of a secondary infection or primary infection of infants born to dengue immune mothers, which led to developing severe disease. This hypothesis is mentioned before, also known as Antibody-Dependent Enhancement (ADE) of the disease [33]. When E-specific antibodies unable to exceed the threshold level to viral opsonization, it enhances the efficiency of the virus to attach and enter into the host cell via FcR-mediated endocytosis [31]. Recently, a study on THP-1 cells infected with the DENV virus shows down regulation of IL-12, IFN-γ, TNF-α and NO and up regulation of IL-6 and IL-10. It signifies the promotion of viral particles through suppression of antiviral immune response via FcR mediated entry. prM antibodies also enhance the infectivity of immature virions but do not affect the number of progeny virions produced per infected cells [34].
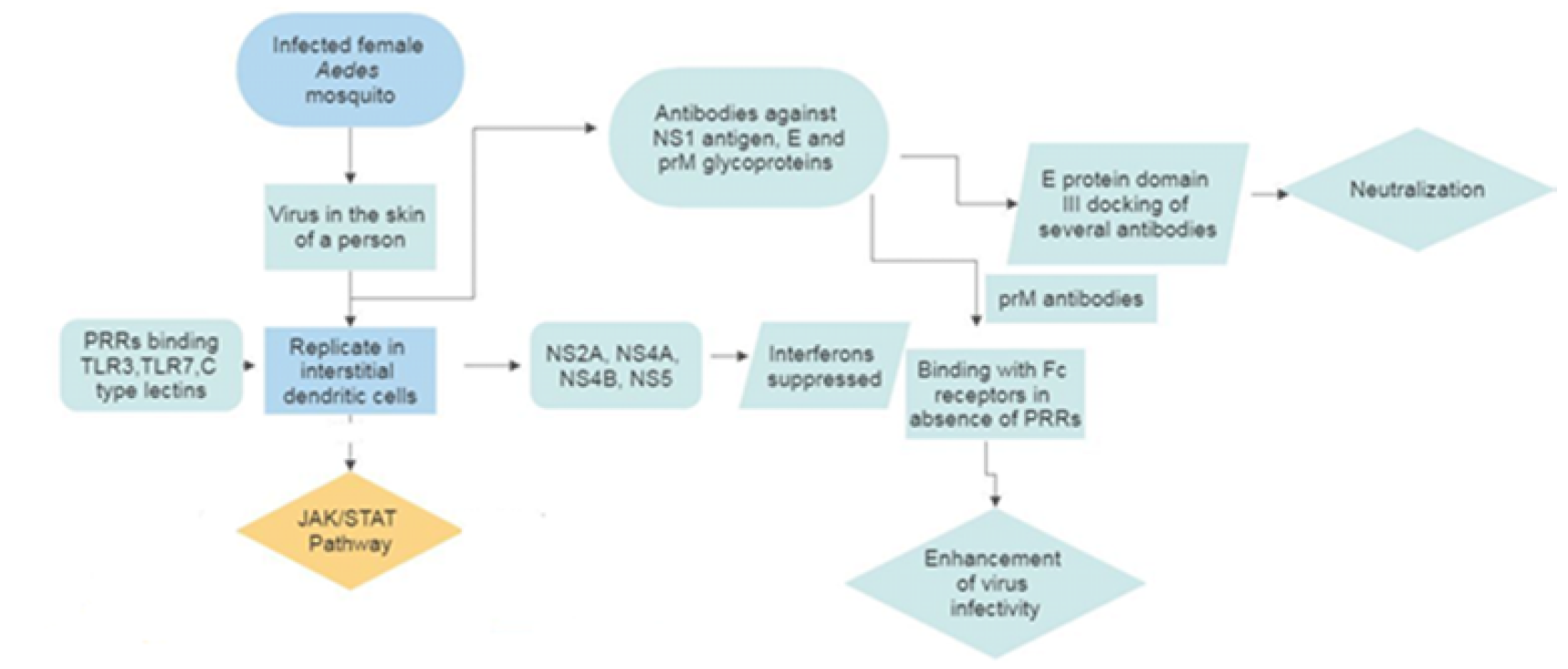
Figure 1: Dengue Virus Replication with Dual Infectivity Outcome
Complexity of ADE
The dengue fever is still endemic in Asia, the Pacific, Africa, the Americas and the Middle East. It is manifested as an asymptomatic or symptomatic disease. There are two types of symptoms: Dengue Fever (DF) and more severe Dengue Hemorrhagic Fever/Dengue Shock Syndrome (DHF/DSS) [35]. It occurs when the antibodies formed from previously infected dengue virus bind with a different serotype of a viral particle. In 1981, the dengue epidemic was reported in Cuba. This DENV 2 epidemic caused more than 300,000 cases in which 10,000 were severe to very severe cases and 158 fatalities including 101 children. But it was found that they were previously infected by DENV 1 virus during the epidemic of 1977-1979. On the other hand, those children born after the 1977 epidemic, showed only primary infection [36]. But almost 20 years after, again DHF/DSS was confirmed in the Cuban epidemics of 1997 (DENV2) and 2001-02 (DENV-3) [37]. In an in vitro study, it was reported that infants who developed DHF after a primary infection had ADE in their mother's sera. This suggests that the maternal antibody to DENV declines constantly through three functional stages: neutralizing, enhancing virus growth and antibody degradation [38]. Therefore, the decrease of cross-reactive neutralizing antibody titers in the serum could be the reason for more severe secondarily infections. Also, DENV 1 followed by DENV 2 and DENV 1 followed by DENV 3 has been reported to associate with greater disease severity [39].
Mechanism of ADE
To neutralize infected DENV particle, an adequate level of high-affinity antibody is required to target the epitopes that located in domain II of the envelope protein. These high-affinity antibodies require poorly accessible epitopes also. But most epitopes are capable to elicit ADE due to low concentration or low-affinity antibody which could poorly access the epitopes to neutralize. For both cases, these sub-neutralizing or non-neutralizing antibodies can redirect the disease in its severe form by causing antibody-dependent enhancement [40]. It has been reported that the virus by its dynamic motion exhibits partially blocked epitopes for antibody binding [41]. When these non/sub neutralizing antibody-virus complexes are internalized in to host cell via FcγRs, it leads to produce higher viral production rather than neutralizing in primary infection. Due to the high number of infected Fcγ-R-bearing cells, it accelerates more internalization and cellular infection in secondary infection [42]. So, it could be hypothesized that patient with high severity has higher viremia compared to others with dengue fever. The molecular mechanism behind ADE is mostly reported in in vitro studies of dengue virus.
Usually, the virus evades early innate immune detection by suppressing NOS2 synthesis followed by down regulation of RIG-1 and MDA-5 protein expression. On the other hand, it up regulates IL-10 and autophagy-related proteins like DAK and Atg5-Atg12. It has been seen DENV 2 infected AG129 mice (IFN-α/β/γ receptor-deficient), born to DENV 1 infected mothers show a high level of disease severity and high level of IL-6 and TNF-α production than those born to naïve mothers [43]. After infection with the virus, it inhibits STAT1 and interferon signaling. Activation of Janus kinase Jak1, Tyk2 and signal transducers of STAT1 and STAT2 up-regulate many genes after a viral infection which stimulates IFN production. This activation is mediated by Type-1 IFN followed by stimulation of IFN receptor subunits IFNAR1 and IFNAR2 [44]. So it is essential to antagonize IFN to maintain a high viral load in the blood. NS5 protein inhibits the IFN signaling by targeting the host protein prolidase. NS5 also degrades STAT2 by engaging the host Ubiquitin-Protein ligase E3 component N-recognin (UBR4) [45].
In other studies, reduction level of Nitric Oxide (NO) production in patients with secondary DENV hemorrhagic fever is reported but not in primary DENV infected patients. In the cell, it is produced by nitric oxide synthase 2 (NOS) which is an integral part of the innate immune system. It helps to inhibit DENV infection by blocking the RNA-dependent RNA polymerase. Evaluating NO level at different intervals, it was observed an increased level of NOS2 production in DENV infected cells. Whereas, decreasing the level of NO production in DENV-ADE infected cells suggests a role of ADE in NO suppression [46].
RIG1 and MDA-5 - these two genes are also inhibited in DENV-ADE infection. The role of these genes is to recognize viral RNA and induce the production of type 1 IFN signaling through Mitochondria Antiviral Protein (MAVS). RIG 1 and MDA 5 inactivates viral particles by recognizing small RNA with 5'cap and long genomic RNA and replication intermediates respectively [47]. But recent studies on Peripheral Blood Mononuclear Cell (PBMC) from patients with secondary DENV hemorrhagic fever show a decreased level of RIG-1, MDA-5 and IFN-β Promoter Stimulator (IPS)-1 comparison with patients with only DENV infection [48].
DENV infection also promotes autophagy production. Autophagy helps the cell to survive during nutrient starvation, infection and stress condition. It encloses cellular organelles in a double membranous vesicle called autophagosomes followed by fusion with lysosomes to form autophagolysosomes, the place of digestion of cellular components. In an Atg5-dependent manner, DENV amplifies and promotes autophagy formation. NS4A in DENV particle promotes autophagy formation in the epithelial cell alone [49].
Factors Associated with ADE
Each DENV consists of different genotypes due to the low fidelity of replication. DENV 2 and DENV 3 genotypes are more associated with dengue hemorrhagic fever [50]. Another example of genotype difference is seen that the south Asian genotype is more virulent than the American DENV caused DF [51]. This is due to the nucleotide difference in prM, E, NS4B, NS5 genes and in the 5’ and 3’ UTRs also. The nucleotide variation at amino acid 390 of E protein is very crucial as it determines host range specificity and virulence [52]. Another study reveals the overall infection rates are higher in Asian lineage than the American lineage for their different ability to infect several populations of A. aegypti [53].
The cytotoxic T cell is another factor in ADE, but whether it protects or enhances the disease is still unknown. After virus infection, this cytotoxic T lymphocyte [CD8+] degrades the infected cells by perforin and granzymes. As these APC cells are one of the main targets of the DENV particle, it induces activation, proliferation and migration to infection sites [54]. The DENV infected cells can then be recognized by CD8+ T cells after forming a complex with HLA class I antigens. Interaction between T cell receptor CD3 and HLA+ antigenic peptide on the surface of target cells helps in this recognition process [55]. Thus activation of memory CD8+ T cells is important in heterologous secondary DENV infection due to the massive production of cytokines and immune modulators that characterizes DHF/DSS [56].
Another characteristic of DHF/DSS is the high production of cytokines. It has been reported that the host cell elicits an abnormal immune response against the virus by producing cytokines and immune modulators i.e IL-1β, IL-2, IL-4, IL-6, IL-7, IL-8, IL-10, TNF-α, MCP-1, IFN-α, IFN-γ, etc. This phenomenon is also known as cytokine storm [57]. It is believed that it may be induced due to 'original antigenic sin' or activation of the high level of cross-reactive low avidity T cells. These T cells exhibit not only altered cytokines production to clear the infection but also cause a massive immune activation [57]. IL-4 helps DENV cells to invade dendritic cells and dermal macrophages. Higher expression of surface receptors facilitates the attachment and entry of the DENV virus [58]. Another TH2 cytokine, IL-13 is also involved in dengue infection. IL10 and THF β are other two factors in which IL10 activates SOCS3 to inhibit signaling pathways induced by IL-6, IL-12, IFN-α, IFN-γ. This allows the virus to escape immune recognition [59]. IL 17 has also been associated with intense acute inflammation and tissue injury leading to vascular permeability [60].
Host genetic factors are also important as difference in disease symptoms is seen in the population. Interestingly black people are less vulnerable than white people due to gene polymorphism like human leukocyte antigens (HLAs) alleles and non-HLA genes [59]. Other host factors like deficiency of glucose 6-phosphate dehydrogenase (G6PD) may lead individuals to develop DHF. Individuals with a chronic disease like diabetes mellitus are also more susceptible to DHF [61].
Conventional and Non-Conventional Treatment of Dengue
According to the Centers for Disease Control and Prevention (CDC), U.S. Department of Health and Human Services; there is no specific medicine to treat DF or DHF [62]. Usually, the treatment is based on the symptoms. Drinking plenty of fluids and antipyretic medicines are the first line of the treatments. To avoid the risk of hemorrhage, it is advised not to take any anti-inflammatory medicines or nonsteroidal drugs [63].
Apart from this, there are some non-conventional treatments like Ayurveda, Siddha and Homoeopathy for treating DF or DHF. It has been suggested that in Ayurveda, papaya has a great effect in increasing the platelet count in dengue patients. But there is some controversy as papaya is not an Indian origin plant, though in Ayurveda, there is a medicine named ‘Eranda Karkati’ (meaning ‘castor cucumber’) which somehow resembles papaya. On the other contradiction, papaya is widely used in Siddha medicine, a traditional South Indian medicine [64]. In homeopathy, Eupatorium perfoliatum is the most common medicine for treating dengue fever. Besides, Phosphorus 30cH and Crotalus horridus 30cH are also used in treatment [65].
Acknowledgment
We thank Dr. Anil Khurana, Director General, Central Council for Research in Homeopathy for his constant support and guidance. Further, we would like to thank the Ministry of AYUSH, Govt. of India for giving us financial support for the study of dengue virus. Besides, we thank all the lab members for their constant support and help for laboratory studies.
Guzmán, M.G. and G. Kourí. “Dengue: An update.” The Lancet Infectious Diseases, vol. 2, no. 1, 2002, pp. 33–42. https://doi.org/10.1016/S1473-3099(01)00171-2
Gubler, D.J. “Epidemic dengue/dengue hemorrhagic fever as a public health, social and economic problem in the 21st century.” Trends in Microbiology, vol. 10, no. 2, 2002, pp. 100–103. https://doi.org/10.1016/S0966-842X(01)02288-0
Dejnirattisai, W. et al. “Cross-reacting antibodies enhance dengue virus infection in humans.” Science, vol. 328, no. 5979, 2010, pp. 745–748. https://doi.org/10.1126/science.1185181
Rodenhuis-Zybert, I.A. et al. “Immature dengue virus: A veiled pathogen?” PLoS Pathogens, vol. 6, no. 1, 2010, e1000718. https://doi.org/10.1371/journal.ppat.1000718
Tuiskunen Bäck, A. and Å. Lundkvist. “Dengue viruses—An overview.” Infection Ecology & Epidemiology, vol. 3, no. 1, 2013, pp. 19839. https://doi.org/10.3402/iee.v3i0.19839
Perera, R. and R.J. Kuhn. “Structural proteomics of dengue virus.” Current Opinion in Microbiology, vol. 11, no. 4, 2008, pp. 369–377. https://doi.org/10.1016/j.mib.2008.06.004
Zidane, N. et al. “Thermodynamic stability of domain iii from the envelope protein of Flavi viruses and its improvement by molecular design.” Protein Engineering, Design & Selection, vol. 26, no. 6, 2013, pp. 389–399. https://doi.org/10.1093/protein/gzt010
Kuhn, R.J. et al. “Structure of dengue virus: Implications for Flavivirus organization, maturation and fusion.” Cell, vol. 108, no. 5, 2002, pp. 717–725. https://doi.org/10.1016/S0092-8674(02)00660-8
Kostyuchenko, V.A. et al. “Immature and mature dengue serotype 1 virus structures provide insight into the maturation process.” Journal of Virology, vol. 87, no. 13, 2013, pp. 7700–7707. https://doi.org/10.1128/JVI.00197-13
Rastogi, M. et al. “Flavivirus NS1: A multifaceted enigmatic viral protein.” Virology Journal, vol. 13, no. 1, 2016, pp. 1–10. https://doi.org/10.1186/s12985-016-0590-7
Libraty, D.H. et al. “High circulating levels of the dengue virus nonstructural protein NS1 early in dengue illness correlate with the development of dengue hemorrhagic fever.” The Journal of Infectious Diseases, vol. 186, no. 8, 2002, pp. 1165–1168. https://doi.org/10.1086/343813
Clum, S. et al. “Cotranslational membrane insertion of the serine proteinase precursor NS2B-NS3 of dengue virus type 2 is required for efficient in vitro processing.” Journal of Biological Chemistry, vol. 272, no. 49, 1997, pp. 30715–30723. https://doi.org/10.1074/jbc.272.49.30715
Niyomrattanakit, P. et al. “Identification of residues in the dengue virus type 2 NS2B cofactor that are critical for NS3 protease activation.” Journal of Virology, vol. 78, no. 24, 2004, pp. 13708–13716. https://doi.org/10.1128/JVI.78.24.13708-13716.2004
Matusan, A.E. et al. “Mutagenesis of the dengue virus type 2 ns3 protein within and outside helicase motifs: Effects on enzyme activity and virus replication.” Journal of Virology, vol. 75, no. 20, 2001, pp. 9633–9643. https://doi.org/10.1128/JVI.75.20.9633-9643.2001
Weidman, M.K. et al. “The interaction of cytoplasmic RNA viruses with the nucleus.” Virus Research, vol. 95, nos. 1–2, 2003, pp. 75–85. https://doi.org/10.1016/S0168-1702(03)00164-3
Umareddy, I. et al. “Dengue Virus NS4B interacts with NS3 and dissociates it from single-stranded RNA.” Journal of General Virology, vol. 87, no. 9, 2006, pp. 2605–2614. https://doi.org/10.1099/vir.0.81844-0
Davidson, A.D. “New insights into flavivirus nonstructural protein 5.” Advances in Virus Research, vol. 74, 2009, pp. 41–101. https://doi.org/10.1016/S0065-3527(09)74002-3
Tay, M.Y.F. et al. “Nuclear localization of dengue virus non-structural protein 5: Protection against all four DENV serotypes by the inhibitor ivermectin.” Antiviral Research, vol. 99, no. 3, 2013, pp. 301–306. https://doi.org/10.1016/j.antiviral.2013.06.002
Holmes, E.C. “Molecular epidemiology and evolution of emerging infectious diseases.” British Medical Bulletin, vol. 54, no. 3, 1998, pp. 533–543. https://doi.org/10.1093/oxfordjournals.bmb.a011708
Fibriansah, G. et al. “Structural changes in dengue virus when exposed to a temperature of 37 °C.” Journal of Virology, vol. 87, no. 13, 2013, pp. 7585–7592. https://doi.org/10.1128/JVI.00757-13
Fibriansah, G. et al. “The development of therapeutic antibodies against dengue virus.” Antiviral Research, vol. 128, 2016, pp. 7–19. https://doi.org/10.1016/j.antiviral.2016.01.002.
Rodenhuis-Zybert, I.A. et al. “Dengue virus life cycle: Viral and host factors modulating infectivity.” Cellular and Molecular Life Sciences, vol. 67, no. 16, 2010, pp. 2773–2786. https://doi.org/10.1007/s00018-010-0357-z.
Clyde, K. et al. “Recent advances in deciphering viral and host determinants of dengue virus replication and pathogenesis.” Journal of Virology, vol. 80, no. 23, 2006, pp. 11418–11431. https://doi.org/10.1128/JVI.01257-06.
Bryant, J.E. et al. “Glycosylation of the dengue 2 virus E Protein at N67 Is Critical for Virus Growth in Vitro but Not for growth in intrathoracically inoculated aedes aegypti mosquitoes.” Virology, vol. 366, no. 2, 2007, pp. 415–423. https://doi.org/10.1016/j.virol.2007.05.007.
Zhang, Y. et al. “Conformational changes of the flavivirus e glycoprotein.” Structure, vol. 12, no. 9, 2004, pp. 1607–1618. https://doi.org/10.1016/j.str.2004.06.019.
Yu, I.M. et al. “Structure of the immature dengue virus at low pH primes proteolytic maturation.” Science, vol. 319, no. 5871, 2008, pp. 1834–1837. https://doi.org/10.1126/science.1153264.
Chen, S.T. et al. “CLEC5A is critical for dengue-virus-induced lethal disease.” Nature, vol. 453, no. 7195, 2008, pp. 672–676. https://doi.org/10.1038/nature07013.
Shresta, S. et al. “Critical roles for both stat1-dependent and stat1-independent pathways in the control of primary dengue virus infection in mice.” The Journal of Immunology, vol. 175, no. 6, 2005, pp. 3946–3954. https://doi.org/10.4049/jimmunol.175.6.3946.
Ashour, J. et al. “NS5 of dengue virus mediates STAT2 binding and degradation.” Journal of Virology, vol. 83, no. 11, 2009, pp. 5408–5418. https://doi.org/10.1128/JVI.02188-08.
Lai, C.Y. et al. “Antibodies to envelope glycoprotein of dengue virus during the natural course of infection are predominantly cross-reactive and recognize epitopes containing highly conserved residues at the fusion loop of domain II.” Journal of Virology, vol. 82, no. 13, 2008, pp. 6631–6643. https://doi.org/10.1128/JVI.00316-08.
Pierson, T.C. et al. “the stoichiometry of antibody-mediated neutralization and enhancement of west nile virus infection.” Cell Host & Microbe, vol. 1, no. 2, 2007, pp. 135–145. https://doi.org/10.1016/j.chom.2007.03.002.
Nelson, S. et al. “Maturation of west nile virus modulates sensitivity to antibody-mediated neutralization.” PLoS Pathogens, vol. 4, no. 5, 2008, e1000060. https://doi.org/10.1371/journal.ppat.1000060.
Halstead, S.B. et al. “Antibody-enhanced dengue virus infection in primate leukocytes.” Nature, vol. 265, no. 5596, 1977, pp. 739–741. https://doi.org/10.1038/265739a0.
Chareonsirisuthigul, T. et al. “Dengue virus (DENV) antibody-dependent enhancement of infection upregulates the production of anti-inflammatory cytokines but suppresses anti-denv free radical and pro-inflammatory cytokine production in THP-1 cells.” Journal of General Virology, vol. 88, no. 2, 2007, pp. 365–375. https://doi.org/10.1099/vir.0.82537-0.
Kouri, G.P. et al. “Dengue haemorrhagic fever/dengue shock syndrome: Lessons from the cuban epidemic, 1981.” Bulletin of the World Health Organization, vol. 67, no. 4, 1989, pp. 375–380.
Gonzales, D. et al. “Classical dengue haemorrhagic fever resulting from two dengue infections spaced 20 years or more: havana, dengue 3 epidemic 2001–2002.” International Journal of Infectious Diseases, vol. 9, 2005, pp. 280–285. https://doi.org/10.1016/j.ijid.2004.07.012.
Kliks, S.C. et al. “Evidence that maternal dengue antibodies are important in the development of dengue hemorrhagic fever in infants.” The American Journal of Tropical Medicine and Hygiene, vol. 38, no. 2, 1988, pp. 411–419. https://doi.org/10.4269/ajtmh.1988.38.411.
Kliks, S.C. et al. “Antibody-dependent enhancement of dengue virus growth in human monocytes as a risk factor for dengue hemorrhagic fever.” The American Journal of Tropical Medicine and Hygiene, vol. 40, no. 4, 1989, pp. 444–451. https://doi.org/10.4269/ajtmh.1989.40.444.
Pierson, T.C. “Modeling antibody-enhanced dengue virus infection and disease in mice: Protection or pathogenesis?” Cell Host & Microbe, vol. 7, no. 2, 2010, pp. 85–86. https://doi.org/10.1016/j.chom.2010.02.004.
Lok, S.M. et al. “Binding of a neutralizing antibody to dengue virus alters the arrangement of surface glycoproteins.” Nature Structural & Molecular Biology, vol. 15, no. 3, 2008, pp. 312–317. https://doi.org/10.1038/nsmb.1382.
Vaughn, D.W. et al. “Dengue Viremia Titer, Antibody Response Pattern and Virus Serotype Correlate with Disease Severity.” The Journal of Infectious Diseases, vol. 181, no. 1, 2000, pp. 2–9. https://doi.org/10.1086/315215.
Ng, J.K.W. et al. “First experimental in vivo model of enhanced dengue disease severity through maternally acquired heterotypic dengue antibodies.” PLoS Pathogens, vol. 10, no. 4, 2014, e1004031. https://doi.org/10.1371/journal.ppat.1004031.
MacMicking, J.D. “Interferon-inducible effector mechanisms in cell-autonomous immunity.” Nature Reviews Immunology, vol. 12, no. 5, 2012, pp. 367–382. https://doi.org/10.1038/nri3210.
Morrison, J. et al. “Dengue virus Co-opts UBR4 to degrade stat2 and antagonize type i interferon signaling.” PLoS Pathogens, vol. 9, no. 3, 2013, e1003265. https://doi.org/10.1371/journal.ppat.1003265.
Takhampunya, R. et al. “Antiviral action of nitric oxide on dengue virus type 2 replication.” Journal of General Virology, vol. 87, no. 10, 2006, pp. 3003–3011. https://doi.org/10.1099/vir.0.81880-0.
Reikine, S. et al. “Pattern recognition and signaling mechanisms of RIG-I and MDA5.” Frontiers in Immunology, vol. 5, 2014, pp. 342. https://doi.org/10.3389/fimmu.2014.00342.
Lin, J.C. et al. “Dengue viral protease interaction with NF-κB inhibitor α/β results in endothelial cell apoptosis and hemorrhage development.” The Journal of Immunology, vol. 193, no. 3, 2014, pp. 1258–1267. https://doi.org/10.4049/jimmunol.1302675.
McLean, J.E. et al. “Flavivirus NS4A-induced autophagy protects cells against death and enhances virus replication.” journal of biological chemistry, vol. 286, no. 25, 2011, pp. 22147–22159. https://doi.org/10.1074/jbc.M110.192500.
Weaver, S.C. et al. “Molecular evolution of dengue viruses: contributions of phylogenetics to understanding the history and epidemiology of the preeminent arboviral disease.” Infection, Genetics and Evolution, vol. 9, no. 4, 2009, pp. 523–540. https://doi.org/10.1016/j.meegid.2009.02.003.
Rico-Hesse, R. et al. “Origins of dengue type 2 viruses associated with increased pathogenicity in the americas.” Virology, vol. 230, no. 2, 1997, pp. 244–251. https://doi.org/10.1006/viro.1997.8504.
Leitmeyer, K.C. et al. “Dengue virus structural differences that correlate with pathogenesis.” Journal of Virology, vol. 73, no. 6, 1999, pp. 4738–4747. https://doi.org/10.1128/JVI.73.6.4738-4747.1999.
Kutubuddin, M. et al. “Recognition of helper T cell epitopes in envelope (E) glycoprotein of japanese encephalitis, west nile and dengue viruses.” Molecular Immunology, vol. 28, nos. 1–2, 1991, pp. 149–154. https://doi.org/10.1016/0161-5890(91)90098-5.
Pang, T. et al. “Of cascades and perfect storms: The immunopathogenesis of dengue haemorrhagic fever–dengue shock syndrome (DHF/DSS).” Immunology and Cell Biology, vol. 85, no. 1, 2007, pp. 43–45. https://doi.org/10.1038/sj.icb.7100008.
Mathew, A. and A.L. Rothman. “Understanding the contribution of cellular immunity to dengue disease pathogenesis.” Immunological Reviews, vol. 225, no. 1, 2008, pp. 300–313. https://doi.org/10.1111/j.1600-065X.2008.00678.x.
Suharti, C. et al. “Cytokine patterns during dengue shock syndrome.” European Cytokine Network, vol. 14, no. 3, 2003, pp. 172–177.
Mangada, M.M. and A.L. Rothman. “Altered cytokine responses of dengue-specific CD4+ T cells to heterologous serotypes.” The Journal of Immunology, vol. 175, no. 4, 2005, pp. 2676–2683. https://doi.org/10.4049/jimmunol.175.4.2676.
Alen, M.M. et al. “Antiviral activity of carbohydrate-binding agents and the role of DC-SIGN in dengue virus infection.” Virology, vol. 387, no. 1, 2009, pp. 67–75. https://doi.org/10.1016/j.virol.2009.01.043.
Flores-Mendoza, L.K. et al. “IL-10 and SOCS3 are predictive biomarkers of dengue hemorrhagic fever.” Mediators of Inflammation, vol. 2017, 2017, article 5197592. https://doi.org/10.1155/2017/5197592.
Chiewsilp, P. et al. “Histocompatibility antigens and dengue hemorrhagic fever.” The American Journal of Tropical Medicine and Hygiene, vol. 30, no. 5, 1981, pp. 1100–1105. https://doi.org/10.4269/ajtmh.1981.30.1100.
Pagliari, C. et al. “Human kidney damage in fatal dengue hemorrhagic fever: Results of glomeruli injury mainly induced by IL17.” Journal of Clinical Virology, vol. 75, 2016, pp. 16–20. https://doi.org/10.1016/j.jcv.2015.12.005.
Bravo, J.R. et al. “Why dengue haemorrhagic fever in cuba? i. individual risk factors for dengue haemorrhagic fever/dengue shock syndrome (DHF/DSS).” Transactions of the Royal Society of Tropical Medicine and Hygiene, vol. 81, no. 5, 1987, pp. 816–820. https://doi.org/10.1016/0035-9203(87)90041-1.
Dengue and Dengue Hemorrhagic Fever. Information for Health Care Practitioners, U.S. Department of Health and Human Services, Centers for Disease Control and Prevention, 2009, https://www.cdc.gov/dengue/resources/denguedhf-information-for-health-care-practitioners_2009.pdf.
Dengue Haemorrhagic Fever: Diagnosis, Treatment, Prevention and Control. 2nd ed., World Health Organization, 1997.
Manohar, P.R. “Papaya, dengue fever and ayurveda.” Ancient Science of Life, vol. 32, no. 3, 2013, pp. 131. https://doi.org/10.4103/0257-7941.122994.
Manchanda, R.K. “Dengue epidemic: What can we offer?” Indian Journal of Research in Homoeopathy, vol. 9, no. 3, 2015, pp. 137–140.
