
+91 6002993949
submission@iarconsortium.org
Open Access
ISSN (Print) : 2708-5090
ISSN (Online) : 2708-5104
Although stomata conductance (Gs) and transpiration (Tr) are important for gas exchange (CO2 and H2O), none of the available studies have considered variation in Gs and Tr between old and young leaves in savanna trees such as (Combretummolle, Piliostigmathonningii and Balanites aegyptiaca). The study aimed at addressing variability in Tr and Gs of three species and their response to changes in canopy microenvironment. Measurements were conducted in Ruma National park. Two treatments and three replications were used in the experiment arranged in a completely randomized design. Results revealed that all the three species recorded highest Tr and Gs in young leaves compared to older leaves. P. thonningii exhibited the highest values followed by C. molle and B. aegyptiaca with the least. Highest Gs of 1.092±0.73 µg cm-2 s-1 was recorded in young leaves of P. thonningii. Similarly, highest mean Tr of 15.73±0.92 µg cm-2 s-1 was recorded in young leaves of P. thonningii followed by C. molle (13.02±1.6 µg cm-2 s-1). There was no significant relationship (p<0.05) between Tr of old leaves of all the species with leaf temperature. B. aegyptiaca, and C. molle exhibited a positive relationship between Tr and VPD in old and young leaves with highest coefficient of determination of R2 = 0.9 obtained in B. aegyptiaca and P. thonningii. Our findings show that estimations of the rate of Tr and Gs is species specific and relies on the microenvironment that the selected leaf is exposed to as well as the age of the leaves. When modeling Tr and Gs from humid tropical savanna trees, generalization of leaf physiological traits is likely to present contradicting results that may underestimate or overestimate their gas exchange attributes.
Woody vegetation within the savannah can have very complex crown structure that can be stratified based on the differences in leaf characteristics such as leaf age and location within the body of the plant. Many features of leaf structure and physiology are known to vary with leaf age [1] while the location of the leaf within the crown can lead to the uneven distribution of climate drivers such as irradiance which contribute to the heterogeneity of leaf behavior in response to internal and external cues [2]. Leaves forming and aging throughout the canopy volume creates an important light gradient from the young to the old leaves that are nested within the vertical light gradient [2-3]. Such differences contribute to the heterogeneity in the features of leaves leading to differences in physiological performance [4-6]. To get an integrated picture of whole-plant water consumption from the responses of single leaves within the canopy, both the age of leaf, leaf inclination in response to incident solar radiation and environmental heterogeneity within the canopies of individual plants should therefore be accounted for [1,7-8]. Furthermore, because of variation in vapour pressure deficit (VPD), the micro-climate inside the canopy influences localized evaporative demand [9]. Most studies have only used leaves on the lighted top canopy however, water relations of these leaves may not accurately reflect tree-scale dynamics [10].
Transpiration and stomatal responses to climate drivers are also non-convergent at the leaf, branch and entire plant scales [10-11]. Different water use and stomatal sensitivity also exist between leaf and branch scale and therefore up scaling leaf-scale data to infer canopy scale water conservation to avoid overestimation of plant response to climate drivers. Such studies are important in order to predict the vulnerability of an ecosystem to climate change [10]. Vertical gradients of irradiance, VPD, air temperature and leaf temperature may all be used to clarify how climate variability and micrometeorological factors influence tree function [12]. These are abiotic factors that influence stomatal conductance (Gs) and consequently transpiration (tr)
Many studies on vegetation in the African savanna have only been exhaustively been determined on the photosynthesis [13-14]. Long term experiments have also been performed and determined on Gs and consequently Tr in several species in african humid savanna vegetation in trees and also shortterm diurnal patterns on for some savanna shrubs. However, estimates in seasonal or annual Gs and tr appear to be available only for some vegetation and measurements are taken from a leaves at a single location within the crown in savanna trees. Furthermore, majority of these estimates are subject to significant uncertainty because they are based on indirect micrometeorological measurements rather than direct plant physiological approaches. Moreover, these estimates were carried with assumption of uniform irradiance penetration within the canopies and considered whole tree water use.
None of the studies considered crown heterogeneity factoring in the age and location of the leaves within the crown.
This study was carried out in Ruma which is a typical moist savanna in Kenya. The savanna ecosystem plays an important role in ecosystem function and structure and provide essential ecosystem services and are currently seen as important targets in climate chance mitigation. However, It is distinguished by spatial and temporal variability, with precipitation fluctuating significantly over time and space. Despite the ubiquitous occurrence of Combretum molle, Balanites aegyptiaca and Piliostigmathonningii species in within Ruma, [15] there is limitation of data on the physiological behavior of the leaves these species in the field on leaves within the heterogeneous crown. This study aimed at comparing leaf Tr and Gs by relating crown characteristics in broad leaved (C. molle and P.thonningii) and narrow leaf-size (B. aegyptiaca) for old and young leaves.
Study site
Ruma National Park (000 36' 28" S & 340 15' 24" E), is located in the old Nyanza province, approximately 140 kilometers south of Kisumu, 23 kilometers south of Homabay Town and 425 kilometers west of Nairobi [16]. A component of the park maintained by the National Youth Services (NYS) serves as a youth training camp and contains around 40 hectares of area. The study site is located in Lambwe Valley between the Kanyamwa Escarpment and the Gwasi Hills (Figure 1). The site receives bimodal annual rainfall with maxima in March and May and between October and December, having an average annual rainfall of 1200 - 1600mm.
The majority of the areas within Ruma National Park, including the study site, are dominated by grassland. Approximately a fifth of the park is an evergreen forest located at the valley's lowest point, forming a distinct flora type [16-17]. The remaining habitat consists of wooded grassland dominated by Balanites aegyptiaca or Acacia drepanolobium, Acacia seyal woodland and bush land. Thermeda triandra is the most dominant grass species in forested grassland, whereas Hyparhenia filipendula dominate in wooded grasslend (Muriuki et al., 2003). Along the slopes there are fertile volcanic soils characterized by rocks and deep slopes where Combretum molle and the broad leaved Pilliostigma thonningii are the most dominant woody species. The park is made up of 68% open or sparsely forested grasslands and 32% forest, forests and thickets. The forested grassland sections are typically under 1 meter thick grass cover, which, while good for herbivores, provides poor grazing habitat for low grazers and increases the risk of fire [16,18].
Figure 1: The Map of Ruma National Park Showing the Study Site
Experimental Design
In this study, three trees (1.3-1.8 m in height and 0.17 m in diameter) of C. molle, B aegyptiaca and P. thonningii were selected. The experiment used a completely randomized design (CRD) with four treatments and 3 replications (trees). The treatments (leaves) from two positions in the canopy: upper part of the tree as young leaves and lower part of the canopy as old leaves. The greatest care was taken in picking leaves with similar positions on comparable twiGs as a means of controlling leaf age, which impacts leaf physiology. The measurements did not include any diseased leaves.
Leaf gas Exchange and Microclimate Measurements
Stomatal conductance and transpiration were measured on a fully expanded trifoliate leaf using a Steady state porometer (LI-1600, Li-Cor, Lincoln, NE, USA) with 2-cm circular aperture cap. Additionally, the porometer recorded the following data: air temperature, relative humidity (RH), leaf temperature (Tleaf) and Global radiation (Gs) on the adaxial leaf surfaces. On every sampled leaf, a porometer measurement took place for 20 to 30 seconds. The temperature inside the leaf cuvette was automatically tracked by sensors installed on and within the porometer leaf chamber. Leaves remained attached to the branch throughout measurements during data collection. Weather permitting, measurements was taken hourly from 08:00 to 18:00. Mean Gs and Tr was calculated for each measurement day considering the species and age of the leaf.
Data analysis
Collected data were analyzed in SPSS software (SPSS 20.0 for Windows, SPSS Inc., Chicago, USA). Regression was carried out to establish the effects of microclimate on Tr and Gs. In case of significant effects, means were compared using least significant difference (LSD) with significance level set at p≤0.05.
Leaf transpiration
Young leaves exhibited higher tr compared to older leaves in all the species (Figure 2). Highest mean Tr were 15.73±0.92 µg cm-2 s-1was recorded in young leaves of P. thonningii followed by C. molle (13.02±1.6 µg cm-2 s-1). B. aegyptiaca had the lowest Tr for both young and old leaves recording10.71±2.93 µg cm-2 s-1 and 8.19±1.27 µg cm-2 s-1, respectively.
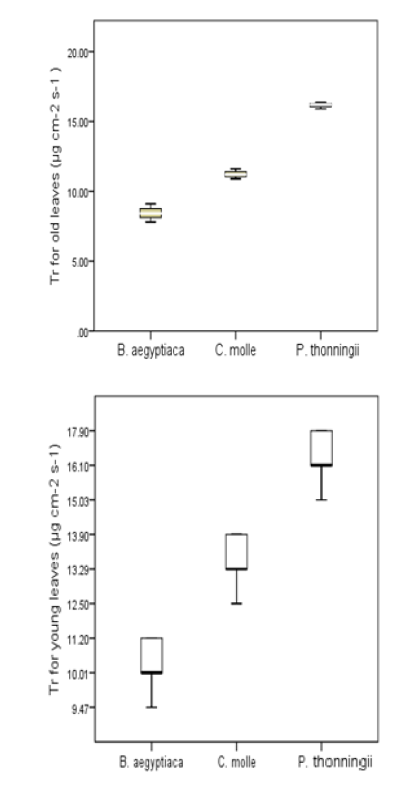
Figure 2: Leaf Transpiration (Tr) (µg Cm-2 S-1) for Old and Young Leaves of C. Molle, B Aegyptiaca and P. Thonningii Tree Species
Stomatal Conductance
Gs followed the same pattern as Tr with highest values of Gs in young leaves 1.092±0.73 µg cm-2 s-1 compared to old leaves 0.64±0.18 µg cm-2 s-1 in P. thonningii (Figure 3). There was a significant variation in Gs for young and old leaves of C. molle but slight differences were observed in B. aegyptiaca. Lowest Gs 0.382±2.54 µg cm-2 s-1 was recorded in old leaves of B. aegyptiaca.
Microclimatic Conditions within the Leaves
Leaf temperature remained slightly constant with fluctuations of ±0.8ºC in all conditions within the canopy. VPD ranged between 2.33-2.70 kPa with slight variation in old and young leaves Table 1. Similarly, solar radiation (Rs) ranged between 237.78 and 795 W/m2 depending on the old or young leaves in all the species. This was similar to RH which was almost similar across the species and canopy location although with variability of 3.32% in young leaves and 2.24% in old leaves.
Relationship between Tr and Gs with Temperature and VPD.
Apart from B. aegyptiaca, all the relationship between Tr and leaf temperature were positive based on regression slopes (b) obtained Table 2. However, there was no significant relationship (p<0.05) between Tr of old leaves of all the species with leaf temperature. C.molle had a higher coefficient of determination of R2 = 0.5 compared to all the other species with R2<0.4. Young leaves of all species had significant (p<0.05) relationship between Tr and leaf temperature.
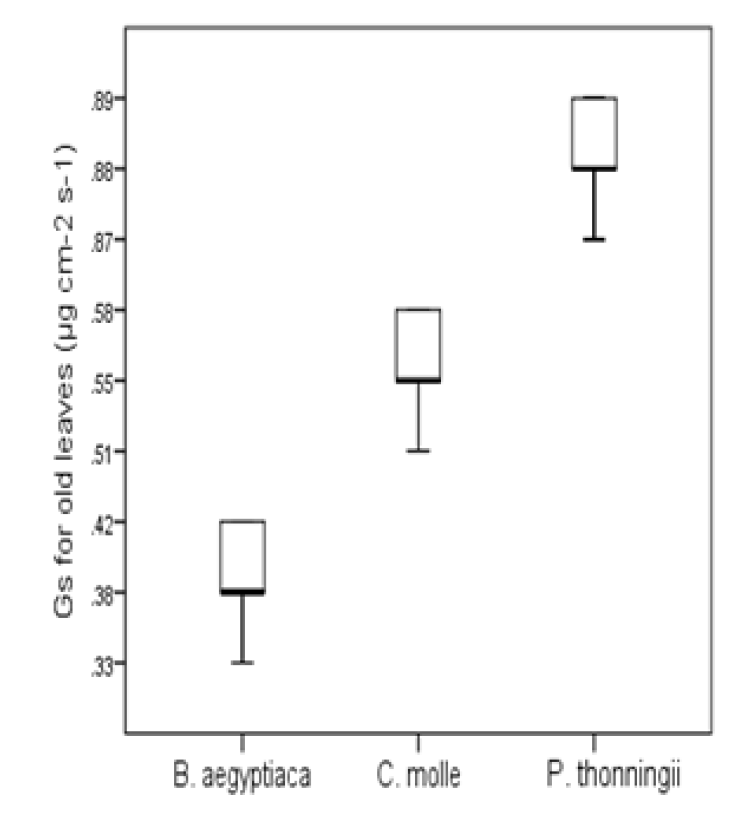
Figure 3: Leaf Stomatal Conductance (Gs) (µg Cm-2 S-1) For Old and Young Leaves of C. Molle, B Aegyptiaca and P. Thonningii Tree Species
Table 1: Measured Microclimate within the Old and Young Leaves of Three Species
Old leaves | ||||||
Species | Rs | Rh |
| Leaf temperature | Vpd |
|
P. thonningii | 354.85 |
| 34.69 | 27.61 |
| 2.41 |
C. molle | 303.25 | 36.21 | 27.50 | 2.33 | ||
B. aegyptiaca | 649.65 | 33.97 | 28.33 | 2.70 | ||
Total | 435.92 | 34.95 | 27.81 | 2.48 | ||
Young Leaves | ||||||
Species | Rs | Rh |
| Leaf temperature | Vpd |
|
P. thonningii | 237.78 |
| 35.07 | 28.04 |
| 2.44 |
C. molle | 581.58 | 36.45 | 28.30 | 2.44 | ||
B. aegyptiaca | 795.37 | 38.39 | 28.01 | 2.50 | ||
Total | 538.25 | 36.64 | 28.12 |
| 2.46 | |
Table 2: Relationship between TR and Leaf Temperature
| Regression between of Tr of old leaves with VPD | ||||
| B. aegyptiaca | B | A | R2 | p- value |
| 0.18 | 26.6 | 0.4 | 0.16 | |
| C. molle | -0.06 | 27.9 | 0.0045 | 0.08 |
| P. thonningii | -1.31 | 48.8 | 0.39 | 0.07 |
| Regression between of Tr of young leaves with VPD | ||||
| B. aegyptiaca | B | a | R2 | p-value |
| 0.18 | 30.2 | 0.8 | 0.05 | |
| C. molle | -0.16 | 30.2 | 0.51 | 0.01 |
| P. thonningii | -0.004 | 28.09 | 0.12 | 0.05 |
Table 3: Relationship between TR and VPD
Regression between of Tr of old leaves with VPD | ||||
B. aegyptiaca | B | A | R2 | p-value |
0.04 | 2.42 | 0.3 | 0.09 | |
C. molle | -0.04 | 2.79 | 0.9 | 0.06 |
P. thonningii | -0.01 | 2.6 | 0.03 | 0.07 |
Regression between of Tr of young leaves with VPD | ||||
B. aegyptiaca | B | a | R2 | p-value |
-0.01 | 2.6 | 0.95 | 0.001 | |
C. molle | 0.01 | 2.25 | 0.36 | 0.05 |
P. thonningii | -0.01 | 2.6 | 0.99 | 0.05 |
As presented in Table 3, B. aegyptiaca and C. molle exhibited a positive relationship between TR and VPD in old and young leaves, respectively. Similar to leaf temperature relationship, there was no significant relationship (p<0.05) between Tr of old leaves of all the species with VPD. Highest coefficient of determination of R2 = 0.9 were obtained in B. aegyptiaca and P. thonningii.
Findings from our study showed the variability in Tr and Gs in young and old leaves. The obtained high Gs in young leaves was attributed to the fact that young leaves tend to have greater stomatal densities in order to maximize photosynthetic exchange and water conductance as shown in other previous studies in different species [5,19-20]. Additionally, variability between young and old leaves may be linked to reports of plants that have developed stomata plasticity pattern by potentially altering stomatal densities to adapt to different environmental stresses within the microclimate to where the leaf resides [21-24]. Zhou et al. [25] reported that old leaves have significantly lower Gs than young leaves as a result of lower stomatal sensitivity to VPD as compared to young leaves. Similar results have been reported by Drake in evergreen conifer stands where stomatal limitations increased with leaf aging. Closing of the stomata with leaf aging can also be due to increase in abscisic acid (ABA) concentration [26-28]. It has been shown that concentration of ABA content increases with increase in age of plant tissues [28], which may have controlled and regulated Tr and Gs in old leaves.
According to Huo and Wang, canopy position and leaf age have a considerable impact on photosynthesis, transpiration rate Tr and specific leaf area (SLA). Similar results have been recorded where Tr in older leaves decreased with leaf age [29]. In this study, the young leaves at the top of the canopy exhibited significantly decreased stomatal density and stomatal index imposed by shady microenvironment from the upper leaves on the lower mature leaves. Similar findings have been reported for tobacco, Vigna sinensis L. and Arabidopsis thaliana. These findings implied that signals that controls stomatal development are conveyed from mature to young leaves in response to variations in irradiance. Similar gas exchange experiments using data collected from leaves at various ages have shown that leaf age is a major contributing factor to gaseous exchange and photosynthetic capacity (Maina and Wa in arid species. The current results add to this growing evidence that Gs and consequently Tr declines as leaves age after full expansion. This behavior is typically associated with the decreased mesophyll conductance and stomatal limitation and typically leaf age affects leaf biochemical properties [1,31]. Our results emphasize canopy resource heterogeneity as a precise factor when assessing photosynthetic performance among leaves at different canopy height. This is because our findings indicate the importance of considering leaf age within the same canopy while estimating physiological properties from plant leaves.
Additionally, since young leaves are typically located at the top of the canopy, the radiative properties of the leaves had greater influence as the leaves tend to heat up causing the intercellular water vapor concentration to increase [9,32]. Heat increase as the sun rises has been shown to increase water vapor concentration and hence an increase in VPD [9]. Additionally, stomatal closure reduces evaporative cooling, which can warm the leaf, increase humidity in the intercellular spaces and consequently raise evaporative demand. Due to the formation of gas bubbles (embolism) in xylem conduits, transport resistance has been shown to increase during soil or atmospheric dryness. The tissues downstream of (distal to) the site where resistance has increased are the only ones affected by this process, which compounds any initial fall in water potential caused by dehydration.
The lack of increase in leaf temperature (Tleaf) between the young and old leaves, where the highest solar radiation was observed, may have been related to an increase in Gs and Tr, which are important in cooling the leaves. Growth light regimes had an impact on the speed and size of the Gs responses to a step shift in PPFD. This may have been a strategy to improve light use efficiency by maintaining open stomata under fluctuating light, reducing the limitation of assimilation by Gs. This may also represent a more conservative strategy in energy under fluctuating light regimes [33]. Other studies have described a reduction in the magnitude of the Gs response through time [34-36]. The control of Gs is predicted to occur at the time when transpiration is peaking and CO2 assimilation is declining and therefore, the ratio of transpiration to assimilation is increasing. Such a shift in control of transpiration, toward the partial control coefficient related to the assimilation rate reflected the fact that the plant leaf system responds in a way that limits the increase in the water loss to carbon gain ratio (i.e. cost/benefit) as unraveled by Maina and Wang in Asian desert species.
Young leaves of all the species exhibited higher Tr and Gs compared to older leaves. This study has revealed that leaf age an important factor in Gs and young leaves responds efficiently to changes in microenvironment compared to older leaves. Our results demonstrate that signals triggered by changes in microenvironment are transferred from older to young leaves and consequently regulate stomatal development. The shade microenvironment may have lowered the stomatal density and stomatal index of the older leaves. Shading of mature leaves may have led to a significant reduction in the epidermal cell density and hence lowered Tr and Gs. Leaves in this microhabitat adapt to light by increasing Tr for leaves that were younger and exposed light. Based on the results, B. aegyptiaca was able to enhance light interception regardless of age of leaves within the canopy but was unable to improve efficiency in resource use by displaying low Tr and Gs. Significant positive relationship in B. aegyptiaca was a response to improved resource investment while P. thonningii and C. molle enhanced their carbon gain through dark reaction, which is favored by low temperatures and hence negative relationship.
Acknowledgement
We acknowledge KWS and NYS for permission to use a section of the Lambwe valley for our experiments. This study was funded by small grants from JOUUST Small Grants for Research (JSGR) 2015/16.
Conflict of Interest
The authors declare no conflict of interest.
Constable, G.A. and H.M. Rawson. “Effect of leaf position, expansion and age on photosynthesis, transpiration and water use efficiency of cotton.” Functional Plant Biology, vol. 7, no. 1, 1980, pp. 89–100.
Durand, M. et al. “Changes in irradiance and vapour pressure deficit under drought induce distinct stomatal dynamics between glasshouse and field-grown poplars.” New Phytologist, vol. 227, no. 2, 2020, pp. 392–406.
Field, C. and H.A. Mooney. “Leaf age and seasonal effects on light, water and nitrogen use efficiency in a California shrub.” Oecologia, vol. 56, nos. 2–3, 1983, pp. 348–355.
Oren, R. et al. “Water balance delineates the soil layer in which moisture affects canopy conductance.” Ecological Applications, vol. 8, no. 4, 1998, pp. 990–1002.
Ewers, B.E. et al. “Effects of stand age and tree species on canopy transpiration and average stomatal conductance of boreal forests.” Plant, Cell & Environment, vol. 28, no. 5, 2005, pp. 660–678.
Bachofen, C. et al. “Light and VPD gradients drive foliar…” (Incomplete reference — title/source missing)
Sobrado, M.A. “Leaf age effects on photosynthetic rate, transpiration rate and nitrogen content in a tropical dry forest.” Physiologia Plantarum, vol. 90, no. 1, 1994, pp. 210–215.
Matloobi, M. “Light harvesting and photosynthesis by the canopy.” Advances in Photosynthesis–Fundamental Aspects, InTech, 2012, pp. 235–256.
Alam, M.S. et al. “A canopy transpiration model based on scaling up stomatal conductance and radiation interception as affected by leaf area index.” Water, vol. 13, no. 3, 2021, pp. 252.
Gu, D. et al. “Non-convergent transpiration and stomatal conductance response of a dominant desert species in Central Asia to climate drivers at leaf, branch and whole plant scales.” Journal of Agricultural Meteorology, vol. 74, no. 1, 2018, pp. 9–17.
Gu, D. et al. “Canopy transpiration and stomatal responses to prolonged drought by a dominant desert species in Central Asia.” Water, vol. 9, no. 6, 2017, pp. 404.
Miller, B.D. et al. “Only sun-lit leaves of the uppermost canopy exceed both air temperature and photosynthetic thermal optima in a wet tropical forest.” Agricultural and Forest Meteorology, vol. 301, 2021, pp. 108347.
Van der Meulen, F. and M.J.A. Werger. “Crown characteristics, leaf size and light throughfall of some savanna trees in southern Africa.” South African Journal of Botany, vol. 3, no. 4, 1984, pp. 208–218.
Walker, B.H. et al. “Size structure analysis of the dominant trees in a South African savanna.” South African Journal of Botany, vol. 52, no. 5, 1986, pp. 397–402.
Ondier, J. et al. “Influence of rainfall amount and livestock grazing on soil respiration in a moist Kenyan savannah.” African Journal of Ecology, vol. 58, no. 1, 2020, pp. 92–99.
Kimanzi, J.K. “Population viability analysis of the endangered roan antelope in Ruma National Park, Kenya and implications for management.” The Scientific World Journal, 2018.
Asila, J. “Ruma National Park animal census research report.” Kenya Wildlife Service, 2004.
Kimanzi, J.K. and C.W. Mbau. “Influence of rainfall fluctuations on population trends of eleven wild herbivores in Soysambu Conservancy, Kenya.” African Journal of Education, Science and Technology, vol. 5, no. 2, 2019, pp. 1–11.
Gonzalez-Dugo, V. et al. “Almond tree canopy temperature reveals intra-crown variability that is water stress-dependent.” Agricultural and Forest Meteorology, vol. 154, 2012, pp. 156–165.
He, L. et al. “Effects of fine-scale soil moisture and canopy heterogeneity on energy and water fluxes in a northern temperate mixed forest.” Agricultural and Forest Meteorology, vol. 184, 2014, pp. 243–256.
Meinzer, F.C. et al. “Control of transpiration from the upper canopy of a tropical forest: The role of stomatal, boundary layer and hydraulic architecture components.” Plant, Cell & Environment, vol. 20, no. 10, 1997, pp. 1242–1252.
Ben-Asher, J. et al. “Irrigation of grapevines with saline water: I. Leaf area index, stomatal conductance, transpiration and photosynthesis.” Agricultural Water Management, vol. 83, nos. 1–2, 2006, pp. 13–21.
Gerardin, T. et al. “Shade and drought growth conditions strongly impact dynamic responses of stomata to variations in irradiance in Nicotiana tabacum.” Environmental and Experimental Botany, vol. 153, 2018, pp. 188–197.
Kromdijk, J. et al. “Predicting light-induced stomatal movements based on the redox state of plastoquinone: Theory and validation.” Photosynthesis Research, vol. 141, no. 1, 2019, pp. 83–97.
Zhou, H. et al. “Leaf-age effects on temperature responses of photosynthesis and respiration of an alpine oak, Quercus aquifolioides, in southwestern China.” Tree Physiology, vol. 35, no. 11, 2015, pp. 1236–1248.
Larbi, A. et al. “Canopy light heterogeneity drives leaf anatomical, eco-physiological and photosynthetic changes in olive trees grown in a high-density plantation.” Photosynthesis Research, vol. 123, no. 2, 2015, pp. 141–155.
Pridgeon, A.J. and A.M. Hetherington. “ABA signalling and metabolism are not essential for dark-induced stomatal closure but affect response speed.” Scientific Reports, vol. 11, 2021, pp. 5751.
Xu, B. et al. “GABA signalling modulates stomatal opening to enhance plant water use efficiency and drought resilience.” Nature Communications, vol. 12, 2021, p. 1952.
Aslam, M. et al. “Effect of leaf age on photosynthesis and transpiration of cassava (Manihot esculenta).” Canadian Journal of Botany, vol. 55, no. 17, 1977, pp. 2288–2295.
Khamis, G. et al. “Morphological and biochemical responses of Balanites aegyptiaca to drought stress and recovery are provenance-dependent.” Journal of Agronomy and Crop Science, vol. 205, no. 5, 2019, pp. 490–507.
Khamis, G. et al. “Morphological and biochemical responses of Balanites aegyptiaca to drought stress and recovery are provenance-dependent.” Journal of Agronomy and Crop Science, vol. 205, no. 5, 2019, pp. 490–507.
Ács, F. “On transpiration and soil moisture content sensitivity to soil hydrophysical data.” Boundary-Layer Meteorology, vol. 115, no. 3, 2005, pp. 473–497.
Harley, P. et al. “Effects of light, temperature and canopy position on net photosynthesis and isoprene emission from sweetgum (Liquidambar styraciflua) leaves.” Tree Physiology, vol. 16, nos. 1–2, 1996, pp. 25–32.
Buckley, T.N. “Modeling stomatal conductance.” Plant Physiology, vol. 174, no. 2, 2017, pp. 572–582.
Li, Y. et al. “Leaf temperature and vapour pressure deficit (VPD) driving stomatal conductance and biochemical processes of leaf photosynthetic rate in a subtropical evergreen coniferous plantation.” Sustainability, vol. 10, no. 11, 2018, p. 4063.
Urban, J. et al. “Stomatal conductance increases with rising temperature.” Plant Signaling & Behavior, vol. 12, no. 8, 2017, e1356534.
