
+91 6002993949
submission@iarconsortium.org
Open Access
ISSN (Print) : 2708-5090
ISSN (Online) : 2708-5104
The distribution patterns and abundance of most pollinator species particularly lepidopterans are poorly documented despite their importance in providing ecosystem services. While these and other organisms are threatened by many aspects of the human enterprise (anthropogenic), climate change may currently be potentially the most severe threat to pollinator biodiversity. Mounting evidence now suggests that there have already been biotic responses to the relatively small climate changes that have occurred this century and butterflies have been depicted as significant indicator species owing to their sensitivity to slightest change in environmental factors. Their abundance in a habitat signals a healthy ecosystem. This study therefore, was conducted in Lochinvar National Park located on the Kafue Flats to collect baseline information about butterfly use of different habitats as indicators of the status of the ecosystem. Data were collected across the three seasons of the year (wet/rainy season-mid November to April; cool dry season-May to August, and hot and dry-mid August to mid-November) by visiting the 16 sites purposely selected because of all year round accessibility and all butterflies sighted on each visit were identified and recorded. A total of 96 butterflies were recorded that belonged to eight families, 37 genera with 57 species. Three families had the largest number of genera and species respectively; Lycaenidae 10 and 12, Pieridae nine and 20, Nymphalidae nine and 12 while the family Danainae had only one genus and one species recorded. Among the sites observed, the Parks offices recorded the highest number of genera and species, 17 and 21 followed by main gate with eight and nine respectively, while the remaining 14 had eight and less. Considering that the study area is in Agro-ecological Region I, it was assumed that the eight families recorded were fairly diverse, but more studies are required to cover a larger area and over a long period of time. Doing so, will enable management to monitor the response of the genera to the impacts of climate change and human disturbance to the butterfly habitats.
Butterflies and moths (Lepidopterans) occur across continents and are a well-known and popular group of insects, enriching the lives of many people. They are a source of food and house hold income in many African countries, possess aesthetical beauty and therefore, an attraction for local and foreign tourists, some are important crop pests, at least in one or two of their life cycles, and also play a critical role in the ecosystem.
In Zambia, several hundred species of tropical and subtropical butterflies occur across the entire country. This great variety of butterflies is largely attributed to the country’s abundant and wide spread water resources, varying habitats which support a wide range of vegetation communities and diversity of plant species. This makes Zambia a suitable location or destination for the study, collection (permitted by DNPW) or simply outdoor pursuit of butterflies and moths alike.
The delightful colour patterns of butterflies confer an inherent aesthetic appeal to both amateur and professional collector and visitor. This is obviously an undeveloped niche in nature based tourism which has the potential to contribute to the increased visitor retention period and satisfaction levels and hence to the growth of the nature based tourism subsector.
[1] however, observed that butterflies among other members of the insect group despite their socio-economic and ecological importance are vulnerable to natural and anthropogenic factors. For instance, in their early stages of life, butterflies have to contend with weather changes, as well as natural enemies, wild fire, habitat loss and in some cases over collection by amateurs and professional collectors, agriculture, expanding industries, use of pesticides and pollution on one hand while the clearing of woody vegetation can reduce food plants (particularly nectar yielding plants) and hence butterfly populations. Without care, quite a number of non-harmful insects can be knowingly or unknowingly be exterminated.
Of critical concern now in the medium to long-term time scales is the survival of butterflies and other insects is the subsisting environmental impacts arising from climate change. Because of their sensitivity to changes in the environment, butterflies have been recognized as a useful biodiversity indicator group of tropical land-use systems because as stated above, they are sensitive and react quickly to subtle changes in environmental and habitat conditions [2-7] Due to their short life cycle, narrow niches and relatively low mobility, they are more sensitive to land-cover and land-use changes than long-lived animals.
It is a known fact that butterfly activities are closely controlled by weather and many species are constrained by climate, usually occupying a small part of the range where host plants as food and cover subsist as acknowledged by [8] Butterflies also play a significant ecological role in agricultural landscapes, are sensitive to changes in the environment, easy to see and study, and for this reason, we considered them to be suitable candidates for monitoring changes in habitat, biodiversity and environmental conditions as earlier reported by [2,9,10] including the impact of landscape and habitat management practices and disturbance regimes in terrestrial ecosystems [11,12]
Despite their smaller size, [13] Schmidt and[14], succinctly reported on their role in the recycling of nutrients (N, P, K) highly needed by crops thereby supporting agriculture (crops). In their larval stage, butterflies feed on leaves of several wild plants found in the agricultural systems and therefore release faeces that contain large amounts of nutrients [15] In addition, butterflies are food to birds and other predators and are hosts to several parasitoids that suppress crop pests [16,17] This important ecological role underscores the need to conserve butterflies in order to sustain the productivity of natural and agricultural landscapes.
In Zambia as is the case with several sub-Saharan African countries, butterflies are not the people’s choice, and most studies as earlier mentioned are skewed towards large and more conspicuous species such as mammals, reptiles and birds. It is indeed common knowledge that monitoring butterflies is not a priority as attention and policy direction of many nature based public institutions as well as NGOs is overwhelmingly tilted towards mega fauna such as elephant, rhino, lion and others. Because of this bias, butterflies remain relatively obscured and little studied, particularly with regard to their ecology, behaviour and functional role in farmland habitats both in natural areas and agricultural landscapes. [18,19] made similar observations that although butterflies made enormous contribution to pollination of plants including crops, there was no marching attention in terms of research in Africa to have a full understanding of their ecology. To the contrary, knowledge of butterflies inhabiting farmland habitats is good in Mediterranean regions compared with the sub-Saharan Africa. The relative scarcity of data on tropical butterfly populations hampers the ability to effectively conserve them, particularly as pollinating agents [20] in agricultural systems.
1.1 Economic Importance of Butterflies and Moths
[21,22] for instance, showed the life cycle of Gonimbrasia belina (Lepidoptera: Saturniidae) and stressed that it was a nutritional food source rich in protein, fatty acids, and minerals and its popularity as food not only for human direct consumption but as a component of livestock feed in the southern region of Africa including Zambia (Figure 1a, b).
Figure 1a: Life cycle of Imbrasia belina a) eggs, b larva –edible stage, c) pupa and d) adult female [22]
Figure 1b: Larva of the popular Imbrasia belina (Source: Favourite snack mopane worm, nydailynews.com)
On the aspect of tourism, butterflies carry an undisputed aesthetic beauty different from large animals particularly in open landscapes such as the Kafue flats where watching butterflies is a priceless adventure.
In conserving butterflies and moths therefore, we need to underscore the concern that climate change poses a huge threat to our butterflies, but that there are also ways to mitigate negative impacts and save them in our gardens, back yards, protected areas and countryside in general [23] Butterflies have short life cycles and thus react quickly to environmental changes. Their limited dispersal ability, larval food plant specialization and close-reliance on the weather and climate make many butterfly species sensitive to fine-scale changes and therefore suitable indicators of habitat condition. Because, butterflies are good biological indicators of environmental change, holistic measures to help them will assist in conserving Zambia’s rich biodiversity. Preserving the natural habitats in areas such as the Lochinvar National Park, which offers an array of large habitats with a mosaic of microclimatic conditions, can undoubtedly help sustain larger and more stable butterfly populations and communities for a much longer time than small areas particularly in isolated urban habitats.
1.2 Impact of climate change on butterflies and the need for their conservation
Across many sub-Saharan African countries, surveys of drivers of butterfly diversity are rare in the literature. In Uganda for instance with a typical wet tropical climate,[24-26,9] showed that there existed no published data describing extensively the diversity of butterflies found in agricultural landscapes in Uganda in relationship to climatic, regional, landscape and local drivers, yet, such information is important for butterfly biodiversity conservation in the rural landscapes. This situation is similar to Zambia where research on butterfly ecology is almost nonexistent. The common literature one is expected to find are field guides such as the publication by [1] a guide to the butterflies of Zambia and to a lesser degree [27] field guide to the butterflies of southern Africa and a handful of other lesser field guides.
Currently, many studies including those by [28-30] show that climatic, regional and landscape factors are important drivers for butterflies populations while other studies also indicate that local factors (availability of nectaring resources) are of foremost importance in explaining the variation in species richness and total density of butterflies as also earlier noted by[31]
Although Uganda where the above studies were carried out is wetter than Zambia, the influence of climatic and anthropogenic factors on butterfly populations is the same. What is critical is to isolate and rate or weigh which local (specific local area), landscape, national and regional climatic factors are important predictors of butterfly species richness and abundance in Zambia and particularly the Kafue Flats where this study was carried out.
1.3 Suitability of butterflies as indicators of climate change
It is also a known fact that insects are particularly useful indicators in the evaluation of landscapes for biological conservation. Among the great diversity of insects, butterflies stand as ideal candidates for ecological study in various landscapes and habitat types. Because of their short life span they quickly respond to environmental variables. For instance, butterflies have been recorded to respond almost instantly to multiple factors such as; plant diversity, habitat complexity, landscape layout, altitude, topography and moisture gradients, and last but not the least climate.
Protected areas such as National Parks and other land management agencies can use butterflies to evaluate the effectiveness of ecosystems based management. Because of their beauty, when they disappear from the landscape, it is relatively easy to notice their absence.
The aim of this study was therefore to assess butterfly taxa in relation to local site characteristics and how they influence diversity in different habitats of Lochinvar National Park, as baseline data upon which medium to long-term monitoring of this ubiquitous but important animals group could be based as one way of securing the biodiversity of the Kafue flats. Patterns of butterfly species occurrence and abundance were analyzed to identify habitats and periods of high butterfly activity, diversity, and uniqueness.
2.1 Study Area Location and Description
2.1.1 Location
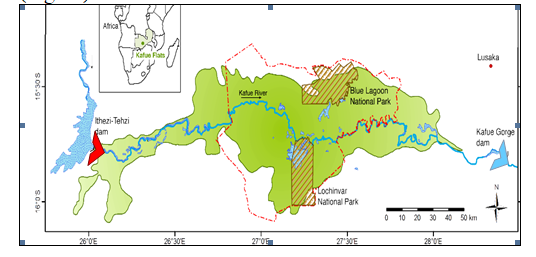
Figure 2 Location of Kafue Flats, habitat for the various species of butterflies
The park is generally flat divided into three major zones, with each zone having a characteristic fauna and flora. The northern third is a flood plain inundated annually by the Kafue River, which also forms the northern boundary. Since 2000, the area has been invaded by alien invasive weed Mimosa pigra. The central zone comprises termitaria grassland, with grassy plains and scattered termite mounds where Euphorbia candelabrum is common. However, since the construction of the Itezhi tezhi dam, there has been woody encroachment in this zone. The southern zone is a woodland area, which is characterized by mainly species of Acacia, Albizia and Combretum. In this zone, are important and popular geological and historical sites. The Gwisho hot springs are about one kilometer from the park headquarters. The Bwanda group of hot springs lies further west and are said to be among the hottest in Zambia. These hot springs are associated with a deposit of gypsum a substance used in the manufacture of cement. The Gwisho hot springs site was historically occupied by groups of late Stone Age people during the third and second millennia BC, while the Sebanzi hill was a site of a more recent Iron Age village that was inhabited for most of the past 1,000 years.
About half of the area is part of the Kafue flood plain making a suitable butterfly habitat. Soils are dark grey of alluvial origin. South of the flood plain, is a flat Terminalia zone on sandy clay to clay soils which are water logged during the wet season while hot springs are indicative of a structural fault which occurs near where the woodland meets the southern boundary of the southern edge of the Terminalia zone. Average annual rainfall is 750mm.
Vegetation is predominantly grass on the flood plain and the most common types are: Oryza birthii, Vossia cuspidata, Echinocloastagnina and Panicum ripens. The commonest herbs are Aeschynomone fluitans and Nymphaea capensis. Steria sphacelata is the characteristic species in the Terminalia grassland. This type of grassland is due to the high water table which is in this zone. South of the National Park is a fire climax woodland of Acacia, Albizia and Combretum spp.
This study was carried out on 16 sites inside Lochinvar National Park. The sites selected were; Hand pump, Parks office, baobab, 400m south of Parks Office, workers compound, Chunga camp, Old lodge, Ecologist's house, main entrance gate, Sebanze hill, airstrip, drum rock, Gwisho road culvert, Chunga lagoon road, Chikolete inside NP, and Munjili area. The sites were located in various habitats representing an array of vegetation communities some affected by various anthropogenic disturbance and management intensities.
The sites selected were accessible all year round. Established monitoring guidelines for butterfly were employed and no specimen was collected except in very special circumstances where the field guides could not positively aid the identification of a specimen. A Field guide to the butterflies of Zambia and the Field Guide to the Butterflies of [27] were used for identification and nomenclature. Meteorology data for each location were obtained from the National Meteorology Agency in Lusaka.
Field sampling of butterflies was conducted by visiting each of the 16 study sites. Taxonomic characteristics were obtained from the field guides. Surveys were conducted in sunny conditions, usually between 09:00- 16:00hrs when butterflies were more active. Counts were made twice per month at 2-week intervals from January to December.
Data Analysis
Descriptive statistics was used to summarize the data and find patterns by obtaining the measures of central tendency such as, mean, median, mode, percentage, frequency and range.
A non-parametric Chi-square goodness of fit test was used to compare observed (fo) and expected (fe) frequencies of the identified parameters.
Specimen collection was only carried out in very exceptional circumstances, particularly when the specimen could not be identified using the field guides. Collection of specimens followed standard protocol.
3.1 Butterfly Abundance and Diversity
In total, 96 observations were made in which eight (8) families, 37 genera, and 61 species respectively were recorded (Table 1 & 2).
Table 1: Butterfly Families recorded in the study area
Family : Papilionidae | |||
Parameter | Details | Parameter | Details |
1) | Graphium konides | 2) | Princeps demodocus demodocus |
Location where sighted | Hand pump | Location where sighted | Hand pump |
Season (Month) | August ( cold and dry season) | Season (Month) | August (cold and dry season) |
|
|
|
|
3)
| Graphium konides | 4) | Graphium angolanus angolanus |
Location where sighted | Hand pump | Location where sighted | Hand pump |
Season (Month) | September (Hot dry season) | Season (Month) | October (Hot dry season) |
|
|
|
|
5)
| Graphium antheus |
|
|
Location where sighted | Hand pump |
|
|
Season (Month) | October (Hot dry season) |
|
|
Family Nymphalidae | |||
1)
| Hamanumida daedalus | 2) | Neptis saclava marpessa |
Location where sighted | Hand pump | Location where sighted | National Park office |
Season (Month) | August | Season (Month) | August |
|
|
|
|
3) | Junonia oenone | 4) | Junonia antilope |
|
|
|
|
Location where sighted | 400m south of Park office | Location where sighted | National Park office |
Season (Month) | July | Season (Month) | June |
|
|
|
|
5)
| Junonia natalica natalica | 6)
| Junonia hierta cebrene |
Location where sighted | Park office | Location where sighted | 400m south of park offices |
Season | June | Season | August |
|
|
|
|
7)
| Junonia artaxia | 8)
| Vanessa cardul |
Location where sighted | 400 south of park offices | Location where sighted | Junior staff compound |
Season (Month) | August | Season (Month) | June |
|
|
|
|
9)
| Byblia navatara acheloia | 10)
| Catacroptera cloanthe cloanthe |
Location where sighted | Park office | Location where sighted | Sebanze hill |
Season (Month) | August | Season (Month) | September |
|
|
|
|
11)
| Charaxes jasius saturnus | 12)
| Hypolimnas misppus |
Location where sighted | Sebanze hill | Location where sighted | Park office |
Season (Month) | September | Season (Month) | December |
|
|
|
|
Family Pieridae | |||
(1)
| Pinacopteryx eriphia eriphia
| 2)
| Eurema spp |
Location where sighted | Park office | Location where sighted | Park office |
Season (Month) | August | Season (Month) | August |
|
|
|
|
3)
| Eurema brigitta brigitta | 4)
| Eronia leda |
Location where sighted | 400 south of park office | Location where sighted | Main gate |
Season (Month) | August | Season (Month) | July |
|
|
|
|
5)
| Mylothris ruppellii rhodesiana | 6)
| Colotis regina |
Location where sighted | Main gate | Location where sighted | Park office |
Season (Month) | July | Season (Month) | May |
|
|
|
|
7)
| Colotis spp | 8)
| Colotis evenina evenina |
Location where sighted | 400m south of park office | Location where sighted | Main gate |
Season (Month) | August | Season (Month) | July |
|
|
|
|
9)
| Colotis pallene | 10)
| Colotis vesta |
Location where sighted | Park office | Location where sighted | Main gate |
Season (Month) | May | Season (Month) | July |
|
|
|
|
11)
Location where sighted | Belenois creona severina
Park office | 12)
| Belenois gidica
Gwisho hot spring |
Season (Month) | June |
| August |
|
|
|
|
13)
| Colotis spp | 14)
| Colotis celimene amina |
Location where sighted - | Park office | Location where sighted - | Sebanze hill |
Season (Month) | September | Season (Month) | September |
|
|
|
|
15)
| Colotis eris eris |
|
|
Location where sighted - | Old Lochnivar lodge |
|
|
Season (Month) | January |
|
|
Family Acraeidae | |||
1)
| Acraea eponina eponina | 2)
| Acraea acrita |
Location where sighted - | Park office | Location where sighted - | Senior staff compound |
Season (Month) | May | Season (Month) | September |
|
|
|
|
3)
| Acraea acara acara | 4)
| Acraea anemosa |
Location where sighted - | Senior staff compound | Location where sighted - | Airstrip |
Season (Month) | October | Season (Month) | December |
|
|
|
|
5)
| Acraea spp | 6)
| Acraea rhodesiana |
Location where sighted - | Drum rock | Location where sighted - | Park office |
Season (Month) | December | Season (Month) | January |
7)
| Acraea eponina eponina | 8)
| Acraea encedon encedon |
Location where sighted - | Park office | Location where sighted - | Park office |
Season (Month) | April | Season (Month) | May |
Family Lycaenidae | |||
1)
| Un identified - 1 | 2)
| Zizula hylax hylax |
Location where sighted - | Junior workers compound | Location where sighted - | Chikolete |
Season (Month) | June | Season (Month) | July |
|
|
|
|
3)
| Unidentified - 2 | 4)
| Tuxentius spp |
Location where sighted - | Gwisho hot spring | Location where sighted - | Park office |
Season (Month) | September | Season (Month) | September |
|
|
|
|
5)
| Axiderses tjoane tjoane | 6)
| Tuxentius calice calice |
Location where sighted - | Park office | Location where sighted - | Main gate |
Season (Month) | September | Season (Month) | September |
|
|
|
|
7)
| Leptotes pirithous pirithous | 8)
| Leptotes pirithous pirithous |
Location where sighted - | Hand pump | Location where sighted - | Hand pump |
Season (Month) | October | Season (Month) | October |
9)
| Unidentified - 3 | 10)
| Axiocerses nyika |
Location where sighted - | Hand pump | Location where sighted - |
|
Season (Month) | October | Season (Month) |
|
|
|
|
|
11) | Lachnocnema bibulus | 12)
| Zizeeria knysna |
Location where sighted - | Old Lochinvar lodge | Location where sighted - | Park office |
Season (Month) | April | Season (Month) | April |
|
|
|
|
13)
| Hypolycaena spp | 14)
| Pinacopteryx eriphia eriphia |
Location where sighted - | Park office | Location where sighted - | Air strip |
Season (Month) | May | Season (Month) | September |
15)
| Catopsila florella |
|
|
Location where sighted - | Hand pump |
|
|
Season (Month) | October |
|
|
Family Hesperiidae | |||
1)
| Sarangesa motozi
| 2)
| Unidentified -1 |
Location where sighted - | Gwisho hotspring | Location where sighted | Senior staff compound |
Season (Month) | September | Season (Month) | November |
3)
| Borbo borbonia borbonia |
|
|
Location where sighted - | Senior staff compound |
|
|
Season (Month) | January |
|
|
Family Satyrinae | |||
1)
| Melantis leda helena | 2)
| Bicyclus anynana centralis |
Location where sighted - | Munjili area | Location where sighted - | Old Lochinvar lodge |
Season (Month) | December | Season (Month) | January |
Family Danainae | |||
1)
| Danaus chrysippus |
|
|
Location where sighted - | Senior staff compound |
|
|
Season (Month) | November |
|
|
Table 2: Number of families and their sightings
No | Family Name | Number of genera | Number of species | Number of months in which recorded | Number of habitats In which recorded | Number of seasons in which recorded | Number of observations |
1 | Papilionidae | 2 | 4 | 3 | 1 | 2 | 5 |
2 | Nymphalidae | 9 | 12 | 5 | 8 | 3 | 21 |
3 | Acraeidae | 1 | 7 | 6 | 6 | 3 | 10 |
4 | Hesperiidae | 3 | 3 | 4 | 3 | 2 | 4 |
5 | Satyrinae | 2 | 2 | 2 | 3 | 1 | 3 |
6 | Pieridae | 9 | 20 | 7 | 9 | 4 | 38 |
7 | Lycaenidae | 10 | 12 | 8 | 7 | 4 | 13 |
8 | Danainae | 1 | 1 | 2 | 2 | 1 | 2 |
Total | 37 | 61 | 37 | 39 | 20 | 96 | |
Lycaenidae, Pieridae and Nymphalidae were the most abundant families and possibly polyphagous in habit, perhaps very well adapted to the environment or may have persisted anthropogenic effects better than others. On the other hand Hesperiidae butterflies being small, dull colored and very active in flight could be difficult to be observed and may have been missed in some instances, and so were recorded the least.
3.2 Family frequency observations
Of the eight (8) families recorded, the most frequently observed were three; Pieridae, Nymphalidae, and Lycaenidae which had the largest number of genera and species respectively (Figure 1). The differences in the number of observations and number of families were found to be significantly different in favour of the three families (χ2 = 37.63, α = 0.05, P < 0.001; χ2 = 84.0, α = 0.05, P < 0.001).
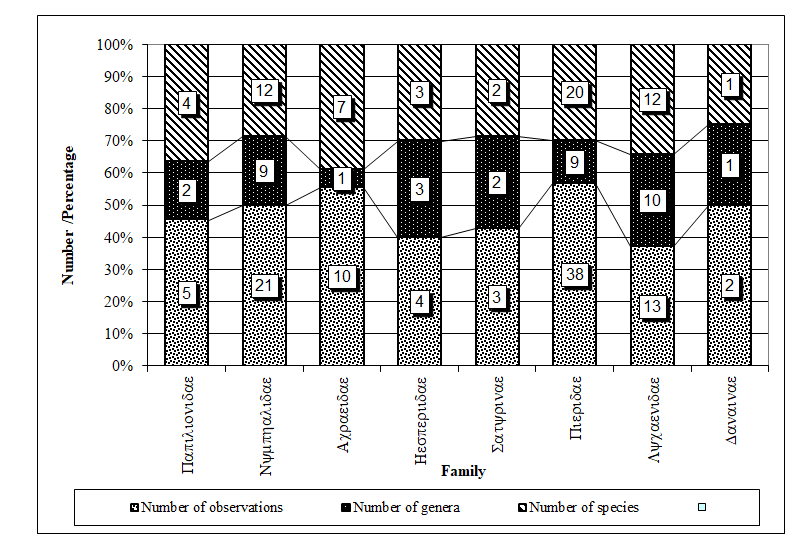
Figure 1: Family frequency observation of families, genera and species
3.3 Uniqueness
The three families were unique in terms of frequency of occurrence between sites. Pieridae 23 % (n = 9), Nymaphalidae 20% (n= 8) and Lycaneidae 18% (n=7) giving a cumulative total of 61% of all observations made between sites. The least were; Papilionidae 2.5% (n=1) observed only in one site and the second was Danainae 5% (n=2) observed only on two sites. No family was found to be a generalist as to be common to all sites.
3.4 Site specific observations
Of the 16 sites visited in this study, three sites accounted for a cumulative total of 51% of species recorded, hand pump 11 % (n = 8), main gate 12% (n = 9) and park office 28% (n = 21). These three sites also had the largest number of genera recorded with a cumulative total of 44% (n = 29), hand pump 6 % (n = 4), main gate 12 % (n = 8) and park office 26% (n = 17). These observations were found to be significantly different (χ2 = 74, α = 0.05, P < 0.001) (Table 3)
Table 3: Study site specific observations of families, genera and species
No | Site | Observations | Families | Genera | Species |
1 | Hand pump | 8 | 2 | 4 | 8 |
2 | Parks office | 34 | 4 | 17 | 21 |
3 | Baobab | 1 | 1 | 1 | 1 |
4 | 400m south of Parks Office | 4 | 2 | 3 | 3 |
5 | Workers compound | 3 | 2 | 3 | 3 |
6 | Chunga camp | 1 | 1 | 1 | 1 |
7 | Old lodge | 6 | 5 | 5 | 5 |
8 | Ecologist's house | 8 | 5 | 6 | 7 |
9 | Main gate | 9 | 4 | 8 | 9 |
10 | Sebanze hill | 4 | 3 | 4 | 4 |
11 | Airstrip | 2 | 2 | 2 | 2 |
12 | Drum rock | 1 | 1 | 1 | 1 |
13 | Gwisho road culvert | 5 | 3 | 4 | 4 |
14 | Chunga lagoon road | 4 | 1 | 4 | 4 |
15 | Chikolete inside NP | 1 | 1 | 1 | 1 |
16 | Munjili area | 1 | 1 | 1 | 1 |
| TOTAL | 82 | 38 | 65 | 75 |
| MEAN | 5 | 2 | 4 | 5(4.6) |
3.5 Comparison across seasons
Most observations of families, genera and species (70%; n = 67) were made in the dry season between May and September when water sources were limited (Figure 4). The month of August had the highest number of observation (22%; n = 21) and the lowest were February and March in which no observations were recorded (Figure 4).
Figure 4: Observation across seasons of the year
4.1 Species abundance
This is the first study to be carried out in this part of Lochinvar National Park, and perhaps the entire Kafue flats area. In that view, therefore, we consider the eight families recorded as a conservatively rich Lepidopteran diversity for a dry savannah habitat such as the Lochinvar National Park which is situated in Agro ecological Region I receiving rainfall not exceeding 700mm per year. [32] in his study of the local habitat characteristics and how they determine butterfly diversity in Kihansi gorge forest of the Udzungwa Mountains in Tanzania recorded only five families with 72 species and considered this as high species richness, although he later acknowledged that the records he obtained in this study were lower than previously published data for this same area in which more than 200 species were recorded. This reduction in species numbers were however attributed to construction and operation of a hydropower facility. [32] he further associated such reduction in species numbers to the
operations of the power plant and the attendant impacts of environmental change recorded in the gorge ecosystem. Based on these impacts on the lepidopteran taxa, they suggested that the differences in butterfly records was due to the disturbances and temporal changes of conditions in their habitats which may have had influenced decline in abundance of species. They also retaliated that there was no study done to date specifically to quantify the impacts of the reduced river flow on the abundance of food resources such as nectar yielding species and availability of suitable breeding cover such as the presence of larval host plant species., the impacts of which have been observed and recorded in other areas as earlier documented by Kitahara, [33]
4.1.2 Importance of water availability
In this study, a preponderance of skewness towards sites with permanent water all year round was observed. There were more and frequent observations in sites with water availability throughout the year than sites without water in the dry season. This is because butterflies have an affinity for water, because they need to take either food or water for their liquid diet. Anthropogenic modifications to the environment such as the provision of water taps played a role in determining butterfly populations distribution and frequency of occurrence.
This explains why most observations were recorded at the Park offices which have water taps that more often than not leak providing a permanent water source and plants growing nearby often being green throughout the year. The hand pump is also a site with a permeant source of water as staff and their families draw water from the hand pump and the overflow often creates a permanent pool of water which the butterflies access. A similar situation obtains at the main gate. The other sites depend on rain water and often remain dry for a large part of the year.
4.1.3 Seasonal Population Dynamics
Seasonal population dynamics was observed as butterfly numbers were more in May-September but less from December to March. The decline in butterfly abundance from December to March may be attributed to the life cycle.
Butterfly and moth taxa have a four (4) life cycle or stages, egg, larva (or caterpillar), pupa and adult. This cycle is called complete metamorphosis. It is likely therefore, that at the beginning and during the rainy season most species would be in their egg, or larva stages, and then towards the end of the rainy season they would be in pupal stage. This may account for the low to no observations in some months of the year as was observed during this study. In this study, the months of February and March had no observations. It is very likely that most genera were in caterpillar stage as this is the time they feed on green vegetable matter and this is also the time that some species cause damage to crops as pests.
genera and species were expected to be recorded as is the case with several other studies in other areas where fewer families were recorded but with a large number of species. Perhaps, savannah conditions of dryness and less cover could be a plausible reason for this, because forested areas often host more butterfly diversity and populations. This assumption is supported by the fact that butterflies are primarily a tropical group of insects and often depend on rain forest plants for larval food. Similarly, according to studies from Manembo - Nembo Wildlife Reserve (Indonesia), more abundant and diversified butterflies were found in riverside habitats in the forest, which is a source of food and provides shelter. In contrast, in Sicily, Italy, grasslands were found to be the most suitable for butterflies because grasslands provide the food and shelter the larvae need.
In this study, the common habitats recorded were open grassland, residential areas, and some bushland and thicket communities which may practically not support high butterfly density compared with forested areas which support butterflies more effectively because of the variety of biomass they provide.
Ecologically, insect species diversity correlates strongly with the structural complexity of habitats and the diversity of vegetation. Food quality also may influences their occurrence. So the, high butterfly diversity reported in forest ecosystems is in response to food quality, as the study by [34] showed that butterflies were not found in disturbed lands; which also supports the idea that degraded habitats affect the population of butterflies. Generally, the diversity of butterflies depends on habitat management and butterfly species richness increases with increasing diversity of plants and decreases with increasing degradation of vegetation.
In this research work therefore, we conclude by suggesting that the results of this study in which we treated the eight families recorded as high diversity should be noted with great care, because, the area has experienced tremendous anthropogenic impacts over the years such as housing, road network construction and others as well as the habitat transformation of the entire Kafue flats after the construction and commissioning of the Itezhi-tezhi hydroelectricity dam in 1979.
The other contemporary factor we should consider is the impact of climate change and its effect on habitat quality in the last few decades. Climate change effects can operate directly through changes in aspects of climate, particularly those that are heat-related, and through changes in habitat diversity. Earlier conceptual models by [35] suggest that regional-scale habitat diversity will decline in a rapidly warming climate and therefore, our remarks on diversity and species richness can only be confirmed with continued monitoring in the future and the continuous review of the isolated records in the past. By reviewing previous records including those relating to temperature rise, it may be possible to estimate previous losses or reduction of some taxa, because physiologically, butterflies and their caterpillars which is the second phase in the process of metamorphosis have an optimum temperature range, within which body processes function optimally. If there are significant local microclimate changes, it may affect their survival and thus have an effect on their numbers and spatial distribution.
We also take note of the influence of the abiotic environment: For instance, drought-periods as well as heavy showers and flooding as has been experienced in the last three years (2019 -2022) in southern Africa affecting mainly Mozambique, Malawi and to a lesser degree Zambia, could alter habitat factors suitable for butterflies. The direct impact would be vegetation structure as it relates to, in some instances nectar yielding plant species which would affect foraging. Therefore, the changing rainfall patterns, increased frequency of droughts we have experienced in the recent past may have had an influence or altered species composition and spatial distribution of some plants and therefore lepidopterans depending on them. If this did occur, then one would assume a low diversity as being the ultimate result.
4.3 Recommendations
In making recommendations for future monitoring and management of lepidopterans, we emphasise the need for the assessment of local drivers influencing richness and abundance of butterflies such as the abundance and distribution of floral nectaring resources among others. Future studies therefore should consider assessing percent cover of flowering plants (trees, shrubs, herbs and weeds), and the number of nectaring plant species. In areas outside the PA it may be worthwhile considering the percentage cover of cultivated floral resources (proportion of cultivated pollinator-dependent and non-pollinator-dependent crops) per unit area.
As regards climate change, cumulative evidence suggests major effects on lepidopterans, leading to species declines and extinctions in some instances as earlier mentioned by [36,23] predicted that the impact of the expected future changes on the lepidopteran populations and distribution will be huge, and action was urgently needed to address this concern. Therefore, measuring the effect of climate change should constitute the first critical step in understanding how butterflies are affected, the speed at which such changes are taking place and the time needed to take amelioration measures. To actualize this on the ground, [37] suggested several methods that could be utilized to study the effect of a changing temperature on lepidopterans. Worth exploring is the method of [38] in which the change of the Community Temperature Index (CTI) proved to be the most sensitive and easy to use indicator for the effect of climate change on butterflies. This is one possibility the management staff of Lochinvar National Park may consider using among other tools.
The park staff should initiate collaboration with butterfly enthusiasts and Nature Based NGOs to work towards promoting awareness among the populations so that land barriers that may hinder mobility across landscapes are minimized and land use practices that mimic agri-environments promoted. The awareness campaigns’ should be widened out to underscore the concern that since lepidopterans are good biological indicators, holistic measures to help them will help conserve Zambia’s rich biodiversity.
Enhance preservation of the natural habitats such as the Lochinvar National Park, which offers a large and diverse set of habitats with a mosaic of microclimatic conditions, can undoubtedly help sustain larger and more stable butterfly populations and communities for a much longer time than small areas particularly in isolated urban habitats. This would also perhaps help facilitate connectivity between areas with large butterfly populations such as protected areas, private land and land under customary tenure where lepidopterans occur in substantial numbers. Greening urban areas where this is feasible should be encourage as this is also likely to achieve similar results.
Away from mega fauna, local scientists should be enticed to supplement the efforts of entomologists who are limited in number to carry out research on butterflies which would in turn provide a frame work to monitor their ecology and to follow changes arising from the impact of climate change and also as a way of building a robust and representative set of indicators.
This baseline study, it is suggested, should be extended to other protected areas so as to further understand the impact and response of lepidopterans to the current anthropogenic activities and climate change.
Author contributions
Both authors contributed equally to conceiving the study, collecting and interpreting data, and developing the final manuscript.
Acknowledgements
We acknowledge the contributions of field staff at Lochnivar National Park who toiled in visiting the 16 sites and many others who critiqued the document from its inception to the final stages.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest. This was a self-funded research activity.
No funding sources
The study was approved by the
Mulungushi university.
Pinhey, E., & Loe, I. (1977). A guide to the butterflies of Zambia. Anglo American Corporation, Central Africa Limited, Lusaka.
Kremen, C.(1994). Biological inventory using target taxa: a case-study of the butterflies of Madagascar. Ecological Applications. 4(3): pp 407– 422.
Libert, M. (1994). Monitoring of numbers and species composition of the butterfly community of two forested hills near Yaounde, Cameroun. Revue d’Ecologie ( La Terre et la Vie). 25(2):pp 151–175
Brown, J.K.S. (1997). Diversity, disturbance, and sustainable use of Neotropical forests: insects as indicators for conservation monitoring. Journal of Insect Conservation. 1(1): pp25–42.
Larsen, T. B. (2008). Forest butterflies in West Africa have resisted extinction … so far (Lepidoptera: Papilionoidea and Hesperioidea). Biodiversity Conservation. 17(12): pp 2833–2847.
Özden, Ö., Ciesla, W. M., Fuller, W. J., & Hodgson, D. J. (2008). Butterfly diversity in Mediterranean islands and in Pentadaktylos Pinus brutia forests of Cyprus. Biodiversity and Conservation, 17(12), 2821-2832.
Pozo, C., Luis-Martínez, A., Llorente-Bousquets, J., Salas-Suárez, N., Maya-Martínez, A., Vargas-Fernández, I., & Warren, A. D. (2008). Seasonality and phenology of the butterflies (Lepidoptera: Papilionoidea and Hesperioidea) of Mexico's Calakmul Region. Florida Entomologist, 407-422.
Fitzherbert, E., Gardner, T., Davenport, T. R., & Caro, T. (2006). Butterfly species richness and abundance in the Katavi ecosystem of western Tanzania. African Journal of Ecology, 44(3), 353-362.
Howard, P. C., Davenport, T. R. B., Kigenyi, F. W., Viskanic, P., Baltzer, M. C., Dickinson, C. J., ... & Mupada, E. (2000). Protected area planning in the tropics: Uganda's national system of forest nature reserves. Conservation biology, 14(3), 858-875.
Cleary, D. F., & Mooers, A. Ø. (2004). Butterfly species richness and community composition in forests affected by ENSO-induced burning and habitat isolation in Borneo. Journal of Tropical Ecology, 20(4), 359-367.
Stork, N. E., Srivastava, D. S., Watt, A. D., & Larsen, T. B. (2003). Butterfly diversity and silvicultural practice in lowland rainforests of Cameroon. Biodiversity & Conservation, 12(3), 387-410.
Öckinger, E., & Smith, H. G. (2008). Do corridors promote dispersal in grassland butterflies and other insects?. Landscape ecology, 23(1), 27-40.
Rogo, L. and Odulaja, A. 2001. Butterflies in two forest fragments at the Kenya coast. African Journal of Ecology. 39(3): -pp 266 –275.
Schmidt, N. B. C., & Roland, J. (2006). Moth diversity in a fragmented habitat: importance of functional groups and landscape scale in the boreal forest. Annals of the Entomological Society of America, 99(6), 1110-1120.
Munyuli, T.M.B. (2010). Pollinator biodiversity and economic value of pollination services in Uganda [PhD dissertation]. [Kampala (Uganda)]: Makerere University. 431 p.
Summerville, K. S., Metzler, E. H., & Crist, T. O. (2001). Diversity of Lepidoptera in Ohio Forests at Local and Regional ScalesHow Heterogeneous is the Fauna?. Annals of the Entomological Society of America, 94(4), 583-591.
Cardoso, P., Scharff, N., Gaspar, C., Henriques, S. S., Carvalho, R., Castro, P. H., & Crespo, L. C. (2008). Rapid biodiversity assessment of spiders (Araneae) using semi‐quantitative sampling: a case study in a Mediterranean forest. Insect Conservation and Diversity, 1(2), 71-84.
Marchiori, O. M., & Romanowski, H. P. (2006). Species composition and diel variation of a butterfly taxocene (Lepidoptera, Papilionoidea and Hesperioidea) in a restinga forest at Itapua State Park, Rio Grande de Sul, Brazil. Rev Bras Zool. 23(2):pp 443–454
Stireman, J. O., Greeney, H. F., & Dyer, L. A. (2009). Species richness and host associations of Lepidoptera-attacking Tachinidae in the northeast Ecuadorian Andes. Journal of Insect Science. 9(39): pp –19.
Bonebrake, T. C., Ponisio, L. C., Boggs, C. L., & Ehrlich, P. R. (2010). More than just indicators: a review of tropical butterfly ecology and conservation. Biological conservation, 143(8), 1831-1841.
Glew, R. H., Jackson, D., Sena, L., VanderJagt, D. J., Pastuszyn, A., & Millson, M. (1999). Gonimbrasia belina (Lepidoptera: Saturniidae): A nutritional food source rich in protein, fatty acids, and minerals. American Entomologist, 45(4), 250-253.
Moyo, S., Masika, P. J., & Muchenje, V. (2019). The potential of Imbrasia belina worm as a poultry and fish feed. A review. J. Anim. Feed Sci, 28(3), 209-219.https://doi.org/10.22358/jafs/112156/2019.
Settele, J., Kudrna, O., Harpke, A., Kühn, I., Swaay, C. van, Verovnik, R., Warren, M., Wiemers, M., Hanspach, J., Hickler, T., Kühn, E., Halder, I. van; Veling, K.., Vliegenthart, A., Wynhoff, I., & Schweiger, O. (2008). Climatic risk atlas of European butterflies. Biorisk 1: pp 1-710
Tumuhimbise, G., Okwakol, M. J. N., & Kangwagye, T. N. (2001). Species diversity of swallowtail butterflies (Papilionidae: Lepidoptera) in North Maramagambo forest. African Journal of Ecology, 39(1), 113-115.
Molleman, F. Kop, A., Brakefield, P.M., Devries, P. J., & Zwaan, B.J. (2006).Vertical and temporal patterns of biodiversity of fruit feeding butterflies in a tropical forest in Uganda. Biodiversity Conservation 15(1): pp 107–121.
Tushabe, H., Kalema, J., Byaruhanga, A., Asasira, J., Ssegawa, P., Balmford, A., & Williams, C. (2006). A nationwide assessment of the biodiversity value of Uganda's important bird areas network. Conservation Biology, 20(1), 85-99.
Migdoll, I. (1992). Field guide to the butterflies of southern Africa. Kyodo Printing Company (Pte) Limited, Singapore.
Chay-Hernández, D. A., Delfín-González, H., & Parra-Tabla, V. (2006). Ichneumonoidea (Hymenoptera) community diversity in an agricultural environment in the state of Yucatan, Mexico. Environmental entomology, 35(5), 1286-1297.
Pickens, B. A., & Root, K. V. (2008). Factors affecting host-plant quality and nectar use for the Karner blue butterfly: implications for oak savanna restoration. Natural Areas Journal 28(3): pp 210–217.
Dover, J., & Settele, J. (2009). The influences of landscape structure on butterfly distribution and movement: a review. Journal of Insect Conservation, 13(1), 3-27.
Pöyry, J., Luoto, M., Heikkinen, R.K., Kuussaari, M. and Saarinen, K. 2009. Species traits explain recent range shifts of Finnish butterflies. Global Change Biology 15: pp 732-743
Rija, A.A. (2022). Local habitat characteristics determine butterfly diversity and community structure in a threatened Kihansi gorge forest, southern Udzungwa Mountains, Tanzania. Springer open Article No.13
Kitahara, M., Yumoto, M., & Kobayashi, T. (2008). Relationship of butterfly diversity with nectar plant species richness in and around the Aokigahara primary woodland of Mount Fuji, central Japan. Biodiversity and conservation, 17(11), 2713-2734.
Rossi, J. P., & Van Halder, I. (2010). Towards indicators of butterfly biodiversity based on a multiscale landscape description. Ecological Indicators, 10(2), 452-458.
Petersm, R. L., & Darling, J. D. S. (1985).The greenhouse effect and nature reserves: Global warming would diminish biological diversity by causing extinctions among reserve species. Biological Science Vol. (35), Issue 11, pp 707 – 717.
Parmesan, C., & Yohe, G. (2003). A globally coherent fingerprint of climate change impacts across natural systems. Nature 421: pp 37– 42.
Van Swaay, C. A. M., Van Strien, A. J., Julliard, R., Schweiger, O., Brereton, T., Heliölä, J., ... & Settele, J. (2008). Developing a methodology for a European butterfly climate change indicator. Report VS2008, 40.
Devictor, V., Julliard, R., Couvet, D., & Jiguet, F. (2008). Birds are tracking climate warming, but not fast enough. Proceedings of the Royal Society B: Biological Sciences, 275(1652), 2743-2748.





























































